Экспериментальная психология
2014. Том 7. № 3. С. 112–125
ISSN: 2072-7593 / 2311-7036 (online)
Анализ поведенческих стереотипов на основе идей колмогоровской сложности: поиск общего методического подхода в этологии и психологии*
Аннотация
Общая информация
* Работа поддержана грантом Российского научного фонда, проект № 14-14-00603.
Ключевые слова: этология, психология, поведенческие стереотипы, охотничье поведение, крысы, теория информации, колмогоровская сложность, сжатие данных
Рубрика издания: Методология психологических исследований
Тип материала: научная статья
Для цитаты: Резникова Ж.И., Пантелеева С.Н., Левенец Я.В. Анализ поведенческих стереотипов на основе идей колмогоровской сложности: поиск общего методического подхода в этологии и психологии // Экспериментальная психология. 2014. Том 7. № 3. С. 112–125.
Полный текст
Постановка задачи
Сравнительная психология в России имеет славное прошлое, восходящее к фундаментальным трудам В. А. Вагнера и Н. Н. Ладыгиной-Котс и их последователей Н. Ю. Войтониса, Н. А. Тих, Г. 3. Рогинского, К. Э. Фабри, Л. А. Фирсова (Мироненко, 2009). Будущее российской сравнительной психологии до сих пор туманно, но в последние годы намечаются перспективы развития этой науки на основе сравнительно-эволюционного подхода (Харитонов, 2013). Актуальной является задача развития методологии, применимой к анализу поведения разных биологических объектов. Данная статья является одним из первых шагов в этом направлении.
Исследование поведенческих стереотипов – область науки, в которой сходятся интересы этологии, поведенческой экологии, психологии и даже психиатрии. Дело в том, что формы поведения, которые для многих видов животных являются нормой, для человека в раннем возрасте могут находиться на грани нормы, а в более зрелом возрасте сигнализировать о патологии. К настоящему времени хорошо известно, что не существует надежных критериев, позволяющих разделить составляющие инстинкта и обучения в поведении животного, не прибегая для этого к специальным экспериментам (Bateson, Gluckman, 2011). Выученные цепочки действий у животных могут стать настолько рутинными и жестко повторяющимися, что по своим проявлениям напоминают цепочки инстинктивных действий (Резникова, Пантелеева, 2005). Еще в начале XX в. психиатры обратили внимание на то, что при некоторых психических заболеваниях люди избирают тактику навязчиво повторяющихся действий, напоминающих ритуализованное поведение животных (Berrios, 2011). Так, одна из характеристик, общая для разных форм аутизма, состоит в крайней ритуализации действий. Психиатры характеризуют такое поведение как стремление к повторению и предохраняющее поведение (Bishop, Richler, Lord, 2006).
Терминология, используемая для описания стереотипов, в психологии носит значительно более тревожный характер, чем в этологии, на каком бы языке ни публиковались исследования. В этологии под поведенческими стереотипами понимают поведенческие последовательности, состоящие из устойчиво повторяющихся элементов (Пантелеева, Данзанов, Резникова, 2010). От стереотипов отличаются стереотипии: повторяющиеся неизменные формы поведения, без очевидной цели и функции (Mason, 1991). Не только у людей, но и у животных стереотипии указывают на патологию в поведении. Они не встречаются у животных в естественной среде и характерны либо для специально выведенных генетических линий (Francis et al., 2003), либо для животных в неволе, причем, они настолько явно свидетельствуют о существенных недостатках в их содержании, что используются как диагностический признак (Broom, Johnson, 1993). Стереотипы, в отличие от стереотипий, не повторяются циклически; поведенческая цепочка имеет начало и завершение, однако, границы между этими формами поведения могут быть и размытыми.
Для задач, которые стоят перед биологами и психологами при описании поведения, необходимо иметь метод для выделения связанных элементов поведения и оценки «меры стереотипности». Выявление граней между стереотипами и стереотипиями важно для определения границ патологии в поведении, как животных, так и людей. Сравнение сложности поведенческих стереотипов может служить мерой для исследования размаха внутри- и межвидовой изменчивости поведения и, соответственно, видовой диагностики. Это особенно важно в зонах возможной гибридизации видов. Для исследования эволюционной преемственности поведения важно не только выделять ключевые элементы и их совпадение в стереотипах разных видов, но и степень их связности. Мы предлагаем в качестве инструмента для количественного описания стереотипов оценку сложности поведенческих последовательностей, основанную на применении идей, связанных с понятием колмогоровской сложности (Колмогоров, 1965). Применение идей теории информации до сих пор ограничивалось исследованием процессов коммуникации (McCowan, Doyle, Hanser, 2002; Oller, Griebe, 2008; Ryabko, Astola, 2006; Ryabko, Reznikova, 2009). Метод, связывающий идеи колмогоровской сложности и математической статистики, имеющий универсальное применение, был разработан лишь недавно (Ryabko et al., 2013).
Поясним основные принципы этого подхода для нашего случая. Если один стереотип состоит из большего числа неповторяющихся элементов, чем другой, то мы можем сказать, что он сложнее, но это не будет полно отражать картину. Например, сравним такие поведенческие цепочки, состоящие из элементов поведения (там же):
АААААААААААААААБААААААААБААААААВААAAAAAАААААААААААГ;
АГБВАББГГВАБББВВГААББВВАГААБВВГАБАБББВВГАБААБАББВВА.
Число элементов в цепочках одинаковое (А, Б, В, Г), последовательность случайная в обеих цепочках, длины цепочек одинаковые, однако вторая, как видно, сложнее. Вышеприведенные рассуждения основаны на неформальном понятии «сложность». Точное определение колмогоровской сложности может быть найдено в работе Колмогорова (Колмогоров, 1965), а здесь мы неформально отметим, что сложность последовательности букв, по Колмогорову – это длина (в двоичном алфавите) минимальной программы, которая «порождает» эту последовательность. В нашем случае алфавит – это весь набор элементарных поведенческих актов, характерных для данного вида, а последовательность букв – демонстрируемое поведение. Поскольку этот подход сложно применить на практике, можно использовать приближение к оценке колмогоровской сложности, которое получают с помощью алгоритмов сжатия данных. Текст, сжатый архиватором, представляет собой некоторую «программу», которая затем при декодировании интерпретируется таким образом, что на выходе мы видим исходный текст. Эта «программа» и является достижимым приближением к колмогоровской минимальной программе (Ryabko, Astola, Gammerman, 2006). Известно, что степень возможного сжатия последовательности символов тесно взаимосвязана с такими ее свойствами как хаотичность и избыточность. В теории информации эти понятия иногда рассматривают в терминах предсказуемости. Так, под хаотичностью подразумевают неопределенность (непредсказуемость) появления в последовательности какого-либо символа из алфавита, а под избыточностью – меру предсказуемости появления последующего элемента, обусловленную предшествующими элементами последовательности. Случайные (непредсказуемые) последовательности символов с низкой избыточностью сжимаются хуже, чем последовательности, обладающие высокой предсказуемостью. На основании этого свойства можно судить об относительной сложности поведения, «записанного» в файлах, допускающих разные степени сжатия архиваторами.
В данном исследовании мы в качестве примера для анализа сложности выбрали поведение очень хорошо изученных животных – лабораторных серых крыс Rattus norvegicus. Их поведение исследовано на разных уровнях интеграции реакций: от отдельных моторных паттернов и их связи с активацией нейронных цепей (Whishaw, Kolb, 2005) до их способности к принятию решений и к сложным формам обучения (Reznikova, 2007). На этой основе мы пытаемся создать экспериментальную модель, применимую к анализу «этологических текстов», генерируемых самыми разными существами, в том числе, и совершенно неизвестными. Представим себе, что мы наблюдаем за поведением инопланетян. Пытаясь его понять, мы сталкиваемся с той же задачей, что и при расшифровке неизвестного нам языка. Для того, чтобы расшифровать «слова», в качестве ключа можно использовать достаточно часто повторяющиеся ситуации, соответствующие определенным сигналам (Резникова, 2008). Для того, чтобы вычленить стереотип из «этологического текста», можно использовать повторяющиеся цепочки элементов и установить их соответствие с конечной «целью», т. е. с завершающим актом. Метод оценки сложности нужен для того, чтобы вычленить из поведенческой цепочки минимальный стереотип, соответствующий ситуации, повторяющейся в жизни живого существа, и отделить его от дополнительных и «шумовых» элементов, которые привносятся гибким, изменчивым поведением.
В статье в качестве первого шага рассматривается охотничье поведение лабораторных серых крыс по отношению к насекомым, выбранное нами как достаточно яркое и выразительное. Мы анализируем наиболее изменчивую часть охотничьего стереотипа, связанную с обнаружением и преследованием жертвы, в двух ситуациях: охоты на подвижную и малоподвижную добычу.
Основные понятия, связанные со стереотипным поведением
К. Лоренц (Lorenz, 1932) впервые выделил видоспецифические модели поведения у птиц. Понятие о видовом (видотипическом) стереотипе поведения было введено А. Н. Промптовым (1940), также на примере птиц, обладающих богатым и демонстративным поведенческим репертуаром. К настоящему времени этологическая литература включает сотни работ, основанных на описании этограмм и выполненных на широком спектре видов, как позвоночных, так и беспозвоночных. Поскольку произошло некоторое размывание и взаимопроникновение терминов и понятий, мы предлагаем, прежде чем анализировать сложность стереотипного поведения, стандартизировать понятия и приемы, применяющиеся при анализе этограмм.
Для описания поведения мы в качестве элементарной единицы выделяем элементарные двигательные акты и позы (Tinbergen, 1951) – для краткости, «элементы поведения». Поведенческой последовательностью мы называем произвольный набор последовательно совершаемых элементов поведения. Примером высоко изменчивой поведенческой последовательности является поисковое поведение у многих видов животных. Устойчиво повторяющиеся «цепочки» элементов поведения выделяем в качестве поведенческих стереотипов. Проявление стереотипов в контексте поведения носит вероятностный характер: отдельные элементы могут выпадать или, наоборот, повторяться, они также могут быть «разбавлены» элементами поведения, не относящимися к данному стереотипу («шумом»). Фиксированным комплексом действий (ФКД) мы, вслед за этологами классической школы, называем генетически обусловленные последовательности поведенческих элементов, постоянные по составу и по порядку совершения (Зорина, Полетаева, Резникова, 1999). Примерами могут служить ФКД захвата добычи, спаривания, чистки тела. Стартуя по принципу «спускового крючка» в ответ на воздействие ключевого стимула, ФКД разворачивается как единая серия действий, вплоть до завершающего акта. ФКД отличается от стереотипа тем, что является неделимым: если старт состоялся, то ФКД дойдет до конца. Так, Лоренц (Lorenz, 1950), в ряду множества других ФКД, приводил в пример охотничье поведение скворца, которое разворачивалось в ответ на ключевой стимул (или даже «внутреннее раздражение») полностью автоматически, пусть и в отсутствие добычи. Возможен был и такой вариант: скворец ловил настоящую муху, но она выскальзывала, и он щелкал клювом вхолостую, а затем даже совершал глотательные движения.
Как стереотип, так и ФКД отличаются от поведенческой последовательности «связностью» своих элементов, т. е. более (для ФКД) или менее (для стереотипа) явно выраженной их взаимозависимостью. Примером стереотипного поведения, включающего в себя ФКД, может служить упорядоченное поведение личинки ручейника Chaetopteryx villosa при строительстве «домика» из песка и других частиц со дна водоема. В процессе строительства прикрепления частиц и продолжительные ощупывания повторяются сериями, одно за другим, во время остановок. Эти серии сменяются сериями повторяющихся отказов от частиц и кратких ощупываний во время передвижения (Непомнящих, 2004).
Как стереотипы, так и ФКД могут быть специфичными для вида. Классическим примером являются устойчивые последовательности элементов поведения при ухаживании и образовании пары у разных видов животных. Отметим, что поведенческие стереотипы могут включать элементы, основанные на приобретенном опыте, столь прочно «встроенные» в поведение индивида, что их трудно отличить от генетически детерминированных (Lorenz, 1950). Если «доводка» стереотипов совершается в контексте однотипных «диалогов» между средой и представителями одного вида, то в итоге получается видотипический стереотип поведения.
Методы и материалы
Мы используем этограммы (т. е. запись поведенческих актов в виде букв некоего алфавита) как пример «биологического текста». В нашем случае алфавит – это весь набор элементарных поведенческих актов, характерный для изучаемых животных, а последовательность букв – демонстрируемое поведение. Мы используем приближение к оценке колмогоровской сложности, которое получают с помощью алгоритмов сжатия данных. Текст, сжатый архиватором, представляет собой некую «программу», которая затем при декодировании интерпретируется таким образом, что на выходе мы видим исходный текст. Эта «программа» и является достижимым приближением к колмогоровской минимальной программе (Рябко, Монарев, 2005; Ryabko et al., 2013). Хотя ранее оценивалась сложность различных, в том числе, и биологических «текстов» (Gusev, Nemytikova, Chuzhanova, 1999), идеи колмогоровской сложности к оценке сложности этограмм были применены лишь недавно (Пантелеева, Данзанов, Резникова, 2010; Reznikova, Panteleeva, Danzanov, 2012). В данной работе мы оцениваем сложность этограмм с помощью метода, разработанного Б.Я. Рябко для анализа биологических текстов на основе идей колмогоровской сложности и проверки статистических гипотез (Ryabko et al., 2013). Этот метод заключается в статистической проверке гипотезы Н0 (последовательности порождаются одним источником), против гипотезы Н1 (последовательности порождаются разными источниками, имеющими разную колмогоровскую сложность). Практически это можно сделать следующим образом: 1) из последовательностей, которые нужно сравнить, выбираются фрагменты (x1...xt) одинаковой длины t так, чтобы к получившимся выборкам можно было применить критерий Манна-Уитни (в каждой из выборок должно быть не менее трех значений признака, в выборочных данных не должно быть совпадающих значений или таких совпадений должно быть очень мало); 2) сложность каждого фрагмента определяется как K(x1...xt ) = |j(x1...xt )| / t, где j – архиватор, а |j(x1 ...xt )| – длина сжатого архиватором фрагмента последовательности; 3) с помощью критерия Манна-Уитни проверяется гипотеза Н0 (между уровнем сложности в рассматриваемых выборках нет существенного различия), против гипотезы Н1 (уровень сложности в рассматриваемых выборках существенно различается).
В данной работе мы применяем предложенный подход для описания охотничьего стереотипа крыс по отношению к подвижной (таракан) и малоподвижной (личинка мучного хрущака) добыче. Известно, что крысы активно охотятся на многих беспозвоночных и мелких позвоночных животных, а их охотничье поведение хорошо изучено (Ivanco, Pellis, Whishaw, 1996). Мы можем предположить, что охота на малоподвижную добычу более стереотипна (и более предсказуема), чем охота на подвижную.
Исследования проводились в 2012–2013 гг., в лабораторных условиях на группе (n=100) беспородных крыс (Rattus norvegicus). Животные содержались по две–три особи одного пола в двухъярусных клетках 45×30×50 см с соблюдением светового режима (8 часов темноты) в зимнее время. Они имели свободный доступ к воде и пище. Для наблюдения за процессом охоты крыс по одной помещали в прозрачную пластиковую арену 45×45×50 см и передерживали в ней в течение 5 мин, после чего предъявляли добычу (бросали в арену сверху) и начинали фиксировать поведение с помощью видеокамеры. Тем особям, которые демонстрировали охотничье поведение в первом же тесте, последовательно предъявляли 3 единицы добычи. Наблюдения длились до завершения поедания добычи, либо, если охота была неуспешной, через 20 мин. Для анализа отбирались только случаи успешной охоты.
В первом эксперименте в качестве подвижной добычи предъявляли имаго мраморного таракана (Nauphoeta cinerea), средняя длина тела 27,93 ±0,22 мм (n=30). Протестировано 81 животное (41 самок, 40 самцов), проведено 15 ч 47 мин видеонаблюдений. Во втором эксперименте предъявляли малоподвижную добычу – личинку мучного хрущака (Tenebrio molitor), средняя длина тела 23,68 ±0,40 мм (n=30). Протестировано 29 животных (22 самки, 7 самцов), проведено 4 ч 43 мин видеонаблюдений. Обработка видеозаписей проводилась посекундно при помощи программы The Observer XT 10.0 (version: 10.1.548 Noldus Information Technology).
Для выделения элементов поведения мы использовали следующий протокол. Для головы (h), передних конечностей (fl), кистей передних конечностей (ha), туловища (с), задних конечностей (hl), челюстей (j) в сочетании с текущим движением (mv) выделены типичные состояния и обозначены числовыми индексами (табл. 1). При помощи этих индексов описывали наблюдаемые элементы поведения, состоящие из блоков локомоций и поз. Всего было выделено 15 элементов поведения (табл. 2). Общая последовательность хищнического поведения крыс, согласно нашим видеозаписям, включает: обнаружение добычи, преследование, укус(ы), манипуляцию с добычей и поедание. Элемент поведения «замирание» (С) использовался в случае, если животное во время охоты прекращало движение и проводило неподвижно более 3 с. Приближение крысы к добыче на расстояние, сопоставимое с размером таракана или мучного хрущака, а также целенаправленное преследование принималось как начало стереотипа. Окончанием полного (завершенного) охотничьего стереотипа считалось поедание добычи. Если крысе не удавалось схватить добычу или добыча вырывалась и убегала, а крыса теряла ее из виду, прекращая искать, констатировался конец незавершенного (неполного) стереотипа. Мы проанализировали только завершенные (полные) охотничьи стереотипы.
Таблица 1. Индексы, используемые при описании элементов поведения крыс
|
Части тела, движения |
Состояние, поза |
|||
|
Голова (h) |
Спокойное, выпрямленное (0) |
Вытянута вперед (1) |
Наклонена вниз (2) |
|
|
Передние конечности (fl) |
Спокойные, касаются земли (0) |
Приподняты перед собой (1) |
Вытянуты (2) |
|
|
Кисти передних конечностей (ha) |
Спокойное, несжатое (0) |
Сжаты в захвате (1) |
|
|
|
Туловище (c) |
Спокойное, выпрямленное (0) |
Вытянуто (1) |
Сгорбленно (2) |
Латерально изогнуто (3) |
|
Задние конечности (hl) |
Спокойные, касаются земли (0) |
Вытянуты, касаются земли (1) |
|
|
|
Челюсти (j) |
Спокойно сомкнуты (0) |
Сжаты в захвате (1) |
|
|
|
Движение (mv) |
Остановка (0) |
Спокойный шаг (1) |
Бег, рысь (2) |
Поворот головы (3) |
|
|
Поворот корпуса на 90° (4) |
Разворот на 180° (5) |
Движение назад (6) |
|
Таблица 2. Элементы поведения в охотничьих стереотипах крыс
|
Символьное обозначение элемента |
Элемент поведения |
Набор индексов |
|
Q |
Бег, рысь |
h-0 fl-0 ha-0 c-0 hl-0 j-0 mv-2 |
|
S |
Спокойный шаг |
h-0 fl-0 ha-0 c-0 hl-0 j-0 mv-1 |
|
W |
Захват добычи зубами |
h-1 fl-0 ha-0 c-1 hl-0 j-1 mv-0,1,2 |
|
E |
Захват добычи передними лапами |
h-0,1 fl-2 ha-1 c-0,1 hl-0 j-0 mv-0,1,2 |
|
R |
Перехват добычи |
h-0,2 fl-1 ha-1 c-2 hl-0 j-0 mv-0 |
|
H |
Откусывание конечностей добычи |
h-2 fl-1 ha-1 c-2 hl-0 j-0,1 mv-0 |
|
G |
Перенос добычи |
h-0 fl-0 ha-0 c-0 hl-0 j-1 mv-1,2 |
|
D |
Принюхивание |
h-0,1 fl-0,1 ha-0 c-0,1 hl-0 j-0 mv-1,2,3 |
|
C |
Замирание |
h-0 fl-0,1 ha-0,1 c-0,2 hl-0 j-0 mv-0 |
|
V |
Поворот корпуса 90° |
h-0,1 fl-0 ha-0 c-3 hl-0 j-0 mv-4 |
|
B |
Разворот корпуса 180° |
h-0,1 fl-0 ha-0 c-3 hl-0 j-0 mv-5 |
|
F |
Поворот головы |
h-0 fl-0,1 ha-0,1 c-0,2 hl-0,1 j-0 mv-0 |
|
Y |
Стойка с поддержкой |
h-0,1 fl-2 ha-0 c-1 hl-1 j-0 mv-0 |
|
U |
Движение назад |
h-0,2 fl-0,1 ha-0 c-2 hl-0 j-0 mv-6 |
|
X |
Чистка |
h-2 fl-1 ha-0 c-2 hl-0 j-0 mv-0 |
Используя полученный «алфавит» из 15 элементов, мы преобразовали отснятый видеоматериал в символьные последовательности, которые перемещали в отдельные текстовые файлы (с расширением txt). Завершенные поведенческие последовательности были объединены и перенесены через пробел в отдельные текстовые файлы. Получено 2 текстовых файла, один из которых содержал завершенные охотничьи стереотипы крыс при охоте на подвижную добычу, второй – завершенные стереотипы при охоте на малоподвижную добычу. Полученные текстовые файлы сжимались при помощи архиватора 7-zip с использованием метода сжатия BZip2, после чего проводилась сравнительная оценка степеней сжатия полных и неполных охотничьих стереотипов. Под степенью сжатия мы понимаем отношение размера заархивированного файла к его исходному размеру. Степень сжатия тем меньше, чем лучше сжимается файл, и наоборот, чем хуже сжимается файл, тем степень сжатия больше. Различия в степени сжатия файлов отражают различия в сложности записанных в них поведенческих последовательностей. Оценивая сложность суммарных файлов последовательностей, необходимо, чтобы все текстовые документы были приведены к одному или максимально близким размерам. Для этого использовался специальный скрипт, который случайным образом удаляет целые (от пробела до пробела) поведенческие последовательности, приводя текстовый файл к определенному размеру. При сокращении из файла исключались стереотипы целиком, а не их части, т.е. итоговый файл составляли только целые стереотипы (как полные, так и неполные).
Для сравнения длин поведенческих последовательностей использовали критерий Стьюдента. Сравнение долей элементов в последовательностях производили при помощи критерия Хи-квадрат.
Результаты экспериментов
В первом эксперименте из 81 протестированной крысы 18 демонстрировали как полные, так и неполные охотничьи стереотипы, 23 – только полные и 13 – только неполные, остальные 27 животных не охотились. Во втором эксперименте из 29 крыс 2 показали наличие как полных, так и неполных охотничьих стереотипов, 21 – наличие только полных, 1 животное – только наличие неполных, и 5 крыс не охотились.
В полных стереотипах имеются различия в частотах встречаемости конкретных элементов (табл. 3).
Таблица 3. Соотношение различных элементов поведения в полных стереотипах
|
Элемент поведения |
Доля элементов в суммарной последовательности, % |
|
|
Охота на таракана |
Охота на личинку мучного хрущака |
|
|
Бег, рысь (Q) |
3,39 |
0 |
|
Спокойный шаг (S) |
3,72 |
17,03 |
|
Захват добычи зубами (W) |
28,36 |
33,34 |
|
Захват добычи передними лапами (E) |
33,03 |
28,26 |
|
Перехват добычи (R) |
15,82 |
3,26 |
|
Откусывание конечностей добычи (H) |
3,39 |
0 |
|
Перенос добычи (G) |
2,33 |
3,62 |
|
Принюхивание (D) |
0,67 |
9,78 |
|
Замирание (C) |
0,26 |
0 |
|
Поворот корпуса 90° (V) |
4,12 |
4,35 |
|
Разворот корпуса 180° (B) |
2,99 |
0 |
|
Поворот головы (F) |
0,47 |
0 |
|
Стойка с опорой на арену (Y) |
0,36 |
0 |
|
Движение назад (U) |
1,02 |
0,36 |
|
Чистка (X) |
0,07 |
0 |
Все выделенные элементы мы разбили на три группы: ключевые (без которых совершение стереотипа невозможно), дополнительные («приготовления» к охоте и поеданию добычи присутствовали не во всех стереотипах) и «шум» (не влияющие на совершение стереотипа: остановки, повороты).
Начинался стереотип с приближения к добыче: бег (Q) или спокойный шаг (S) (в тех редких случаях, когда добыча сама приближалась к крысе слишком близко, охотничий стереотип мог начинаться без фазы преследования), затем следовало принюхивание (D), далее следовал захват добычи зубами (W). За ними следовал захват добычи передними лапами (E), после чего в случае охоты на малоподвижную добычу крыса приступала к поеданию добычи. В случае с подвижной добычей появлялся ряд дополнительных элементов: перехват добычи передними лапами (R), откусывание конечностей насекомого (H), за которым, как правило, следовал очередной перехват (R) и поедание добычи. Элементы следовали в следующем порядке: в случае малоподвижной добычи S-[D]-W-[G]-E, а в случае подвижной Q/S-[D]-W-[G]-E[R]-[H]. В квадратных скобках указаны дополнительные элементы. Отдельные элементы (и блоки элементов), входящие в завершенный стереотип, могут повторяться. Например, повторяются захват добычи зубами с последующим захватом добычи лапами. Отметим, что подобная связка встречается часто и, как можно полагать, представляет ФКД.
Между ключевыми элементами могут быть необязательные составляющие («шум») – например, повороты и остановки. При этом принципиальная схема стереотипа не меняется (рис. 1).
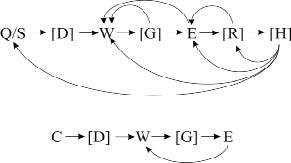
Рис. 1. Принципиальная схема охотничьего стереотипа крыс при охоте на подвижную (вверху) и неподвижную (внизу) добычу. Стрелками показаны переходы между элементами. В квадратных скобках указаны необязательные элементы и шум Длина полных стереотипов у крыс, охотившихся на таракана, варьировала от 2 до 88 элементов (в среднем 22,08 ±1,67), а у охотившихся на личинку мучного хрущака от 2 до 13 (в среднем 4,04 ±0,23). Полные охотничьи стереотипы крыс при охоте на таракана достоверно длиннее стереотипов при охоте на личинку мучного хрущака (p<0,01; t=8,052). Поведенческие последовательности при охоте на личинку мучного хрущака содержали значительно больше ключевых элементов, чем при охоте на таракана (рис. 2) (χ2=12,103; p<0,005).
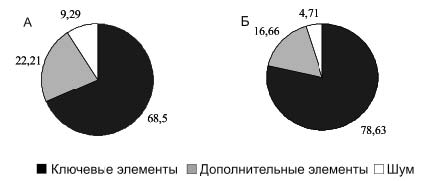
Рис. 2. Доли элементов в охотничьих стереотипах крыс при охоте на подвижную (А) и малоподвижную (Б) добычу
Из двух полученных совокупностей поведенческих последовательностей были сформированы по пять текстовых файлов, которые содержали полные охотничьи стереотипы крыс. Полученные файлы сжимались архиватором, после чего вычислялись степени сжатия (табл. 4). Текстовые файлы, содержащие охотничьи стереотипы крыс при охоте на малоподвижную добычу, сжимаются лучше, чем файлы, содержащие стереотипы при охоте на подвижную добычу (Uэмп=0, Uкрит=1 p≤0,01).
Таблица 4. Степени сжатия случайно сформированных файлов, содержащих фрагменты последовательностей
|
Показатель |
Размер файла до сжатия, байт |
Размер файла после сжатия, байт |
Степень сжатия, % |
|
Текстовые файлы, содержащие случайные фрагменты полных охотничьих стереотипов при охоте на таракана |
248 |
147 |
0,5927 |
|
249 |
146 |
0,5863 |
|
|
250 |
152 |
0,608 |
|
|
250 |
151 |
0,604 |
|
|
249 |
151 |
0,6064 |
|
|
Текстовый файл, содержащий случайные фрагменты полных охотничьих стереотипов при охоте на личинку мучного хрущака |
250 |
108 |
0,432 |
|
250 |
101 |
0,404 |
|
|
249 |
110 |
0,4418 |
|
|
249 |
111 |
0,4458 |
|
|
248 |
107 |
0,4315 |
Заключение
Таким образом, мы, в качестве первого шага к анализу стереотипного поведения в широком контексте этологических и психологических задач, создали экспериментальную модель, применив новый метод оценки сложности поведенческих «текстов» (Ryabko et al., 2013) к достаточно выразительному хищническому поведению хорошо изученного вида грызунов. Выявив «словари» для двух типов такого поведения у крыс – при их охоте на подвижную и малоподвижную добычу – мы затем на большом видеоматериале применили сжатие двух типов «текстов» с помощью архиваторов и сравнили их сложность. Это дало возможность вычленить из поведенческой цепочки базовый минимальный стереотип и отделить его от дополнительных и «шумовых» элементов, которые привносятся гибким, изменчивым поведением. В нашем случае оказалось, что поведенческие стереотипы у крыс при охоте на подвижную добычу характеризуются большей сложностью по сравнению со стереотипами при охоте на малоподвижную добычу. Они значительно длиннее и содержат меньше ключевых элементов и больше дополнительных и «шумовых», т. е. необязательных для завершения стереотипа и привнесенных гибким изменчивым поведением.
Исследуя хищническое поведение крыс, мы использовали цепочки элементов, естественным образом повторяющиеся в жизни живого существа, и установили их соответствие с конечной «целью», т. е. с завершающим актом. Сопоставление сложности стереотипов крыс при охоте на подвижную и малоподвижную добычу показало хорошую применимость метода анализа поведенческих «текстов», основанного на идеях колмогоровской сложности, к выявлению закономерностей стереотипного поведения и его перспективность для формирования общего методического подхода в этологии и в психологии.
Литература
- Зорина З.А., Полетаева И.И., Резникова Ж.И. Основы этологии и генетики поведения. М.: МГУ, 1999. 383 c.
- Колмогоров А.Н. Три подхода к определению количества информации // Проблемы передачи информации. 1965. Т. 1. № 1. С. 3–11.
- Мироненко И.А. О прошлом, настоящем и будущем российской сравнительной психологии // Методология и история психологии. 2009. Т. 4. Вып. 2. С. 45–58.
- Непомнящих В.А. Как животные решают плохо формализуемые задачи поиска // Синергетика и психология: Тексты. Выпуск 3: Когнитивные процессы / Ред. Аршинов В.И., Трофимова И.Н., Шендяпин В.А. М.: Когито-Центр, 2004. C. 197–209.
- Пантелеева С.Н., Данзанов Ж.А., Резникова Ж.И. Оценка сложности поведенческих стереотипов у муравьев на примере анализа охотничьего поведения Myrmica rubra (Hymenoptera, Formicidae) // Зоологический журнал. 2010. Т. 89. № 12. С. 500–509.
- Промптов А. Н. Видовой стереотип поведения и его формирование у диких птиц // Докл. АН СССР. 1940. Т. 27. № 2. С. 171–175.
- Резникова Ж.И. Когнитивное поведение животных, его адаптационная функция и закономерности формирования // Вестник НГУ. Серия: Психология. 2009. Т. 3. № 2. C. 53–68.
- Резникова Ж.И. Современные подходы к изучению языкового поведения животных // Сборник "Разумное поведение и язык. Коммуникативные системы животных и язык человека". М.: "Языки славянских культур", 2008. С. 293–337.
- Резникова Ж.И., Пантелеева С.Н. Экспериментальное исследование формирования охотничьего поведения в онтогенезе муравьев // Доклады Академии наук. 2005. Т. 401. № 1. С. 139–141.
- Рябко Б.Я., Монарев В.А. Экспериментальное исследование методов прогнозирования, базирующихся на алгоритмах сжатия данных // Проблемы передачи информации. 2005. Т. 41. № 1. С. 75–78.
- Харитонов А.Н. Эволюционная и сравнительная психология в России: традиции и перспективы // М.: Институт психологии РАН. 2013. 432 с.
- Bateson P., Gluckman P. Plasticity, Robustness, Development and Evolution. Cambridge: Cambridge University Press, 2011. 166 p.
- Berrios G.E. Eugen Bleuler’s Place in the History of Psychiatry // Schizophrenia Bulletin. 2011. Vol. 37. P. 1095–1098.
- Bishop S.L., Richler J., Lord C. Association between restricted and repetitive behaviors and nonverbal IQ in children with autism spectrum disorders // Child Neuropsychol. 2006. Vol. 12. P. 247–267.
- Broom D.M., Johnson K.G. Stress and Animal Welfare. Dordrecht: Kluwer (formerly Chapman and Hall), 1993. 211 p.
- Francis D.D., Szegda K., Campbell G., Martin W.D., Inse T.R. Epigenetic sources of behavioral differences in mice // Nature Neuroscience. 2003. Vol. 6. P. 445–446.
- Gusev V.D., Nemytikova L.A., Chuzhanova N.A. On the complexity measures of genetic sequences // Bioinformatics. 1999. Vol. 15. № 12. P. 994–999.
- Ivanco T.L., Pellis S.M., Whishaw I.Q. Skilled forelimb movements in prey catching and in reaching by rats (Rattus norvegicus) and opossums (Monodelphis domestica): relations to anatomical differences in motor systems // Behavioural Brain Research. 1996. Vol. 79. № 1. P. 163–181. DOI 10.1016/0166-4328(96)00011-3.
- Lorenz K. Betrachtungen über das Erkennen derarteigenen Triebhandlungen der Vögel // J. für Ornithologie. 1932. Vol. 80. P. 50–98.
- Lorenz K. The comparative method in studying innate behaviour patterns // Symposium of the Society of Experimental Biology. 1950. Vol. 4. P. 221–268.
- Mason G.J. Stereotypies: a critical review // Animal Behaviour. 1991. Vol. 41. P. 1015–1037.
- McCowan B., Doyle L.R., Hanser S.F. Using information theory to assess the diversity, complexity, and development of communicative repertoires // Journal of Comparative Psychology. 2002. Vol. 116. № 2. P. 166–172.
- Oller D.K., Griebe U. Evolution of Communicative Flexibility: Complexity, Creativity, and Adaptability in Human and Animal Communication. Cambridge: MIT Press, 2008. P. 305–325.
- Reznikova Zh. I. Dialog with black box: using Information Theory to study animal language behaviour // Acta ethologica. 2007. Vol. 10. № 1. P. 1–12.
- Reznikova Zh., Panteleeva S., Danzanov Zh. A new method for evaluating the complexity of animal behavioral patterns based on the notion of Kolmogorov complexity, with ants' hunting behavior as an example // Neurocomputing. 2012. Vol. 84. P. 58–64. DOI 10.1016/j.neucom.2011.12.019.
- Ryabko B., Astola J., Gammerman A. Application of Kolmogorov complexity and universal codes to identity testing and nonparametric testing of serial independence for time series // Theoretical Computer Science. 2006. Vol. 359. P. 440–448.
- Ryabko B., Reznikova Zh. The use of ideas of information theory for studying “language” and intelligence in ants // Entropy. 2009. Vol. 11. P. 836–853.
- Ryabko B., Reznikova Zh., Druzyaka A., Panteleeva S. Using Kolmogorov complexity for studying biological texts // Theory of Computing Systems. 2013. Vol. 52. № 1. P. 1–17. DOI 10.1007/s00224-012-9403-6.
- Tinbergen N. The Study of Instinct. London: Oxford University Press, 1951. 228 p.
- Whishaw I.Q., Kolb B. The Behavior of the Laboratory Rat: A Handbook with Tests. Oxford, England: Oxford University Press, 2005. 560 p.
Информация об авторах
Метрики
Просмотров
Всего: 5940
В прошлом месяце: 35
В текущем месяце: 42
Скачиваний
Всего: 996
В прошлом месяце: 3
В текущем месяце: 1