Экспериментальная психология
2015. Том 8. № 1. С. 73–84
ISSN: 2072-7593 / 2311-7036 (online)
Ранний сенсорный опыт определяет разнообразие исследовательского поведения в зрелом возрасте*
Аннотация
Общая информация
* Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проект № 13-04-00741 «Измененное поведение матери – причина патологии потомства»).
Ключевые слова: онтогенез поведения крыс, «открытое поле», удаление вибрисс, скорость движения, ускорение движения, исследовательская активность
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
Для цитаты: Шишелова А.Ю., Алиев Р.Р., Раевский В.В. Ранний сенсорный опыт определяет разнообразие исследовательского поведения в зрелом возрасте // Экспериментальная психология. 2015. Том 8. № 1. С. 73–84.
Полный текст
Введение
Процессы, происходящие в развивающейся нервной системе во время раннего постнатального онтогенеза, являются основой для формирования сложных видов целенаправленного поведения, используемых во взрослом возрасте. Функциональные системы ранних поведенческих актов в последующем подвергаются реорганизации в результате взаимодействия особи со средой, расширения спектра потребностей и получения нового опыта в процессе их удовлетворения. По мере усложнения поведения и обучения в мозге происходит формирование новых специализаций нейронов и аккомодационная реконсолидация уже существующих нейронных взаимодействий (Сварник и др., 2011; Созинов и др., 2013).
В раннем онтогенезе развитие поведения и психики во многом определяется характеристиками поступающих от ближайшего социального окружения видоспецифических сигналов (Ardiel, Rankin, 2010; Curley et al., 2009; Curley et al., 2011; denenberg, 2000; Pryce, Feldon, 2003; Rodel, Meyer, 2011). У незрелорождающихся животных в первые недели жизни основным источником видоспецифической афферентации являются мать и сибсы (Ardiel, Rankin, 2010; Denenberg, 2000; Pryce, Feldon, 2003). Сенсорную информацию о них норковые животные получают главным образом с помощью вибрисс, преимущественно усовых (mystacial) (Grant, Mitchinson et al., 2012; Grant, Sperber et al., 2012; Landers, Sullivan, 2012; Miyazaki et al., 2012).
Созревание вибриссной системы у крыс происходит постепенно в течение первых 3-х недель жизни (Grant, Mitchinson et al., 2012; Erzurumlu, 2010; Shoykhet, Simons, 2008). Первым этапом является формирование корковых проекций усовых вибрисс в виде топически организованных скоплений нейронов – «бочонков». Этот процесс у крысят завершается к 5-му – 7-му дню постнатального онтогенеза (Erzurumlu, 2010) и позволяет оценивать количественные и качественные характеристики предметной среды. Однако в этот период восприятие через вибриссы является «пассивным» и осуществляется благодаря случайным отклонениям вибрисс при контактах с сибсами и матерью (Grant, Mitchinson et al., 2012). Это согласуется с характером поведения крысят в этом возрасте: большую часть времени они спят, прижавшись друг к другу, ходьба, стойки и другие, характерные для взрослых животных, проявления двигательной и исследовательской активности отсутствуют (Bolles, Woods, 1964).
После 8-го дня жизни начинают появляться активные движения усовых вибрисс («whisking»). К концу 2-й недели, перед открытием глаз, движения вибрисс у крысят становятся целенаправленными, «ощупывающими», сходными по характеру с наблюдающимися у взрослых особей. С этого возраста вибриссная система начинает активно участвовать в контроле текущего поведения (Grant, Mitchinson et al., 2012; Erzurumlu, 2010). В это же время формируются основные индивидуальные поведенческие акты: ходьба, манипуляционная активность, груминг, стойки с опорой и без опоры (Bolles, Woods, 1964; Grant, Mitchinson et al., 2012).
Для формирования ранних поведенческих реакций и открытия глаз характерным является наличие высокой степени корреляций между сроками их появления (Шишелова, Раевский, 2009). Согласно предложенной нами гипотезе, такая синхронность является фактором, обеспечивающим интеграцию разрозненно сформированных ранних форм поведения в новые функциональные системы, определяющие многообразие и, возможно, индивидуальный характер поведения. Ранее нами было показано, что проявление этой синхронности зависит от видоспецифической афферентации в раннем онтогенезе (Шишелова, Раевский, 2009). Ограничение ее за счет выстригания вибрисс с 9-го по 20-й дни жизни разобщает созревание указанных форм поведения. Последнее позволило сформулировать задачу настоящей работы – определение качества и степени влияния раннего сенсорного опыта на многообразие и индивидуальные характеристики исследовательской активности – одного из важнейших для адаптации видов поведения грызунов.
Методика
Работа выполнена на выращенных в лаборатории самцах крыс линии Вистар, полученных от пяти первородящих самок. Все эксперименты были выполнены в соответствии с требованиями Директивы Совета Европейского сообщества (86/609/ЕЕС) об использовании животных для экспериментальных исследований.
На 2–й день после рождения проводили осмотр пометов, в каждом оставляли по 8 крысят. С 9–го по 20–й день жизни у половины крысят из каждого помета ежедневно ножницами выстригали усовые вибриссы (опытная группа), у другой половины помета теми же ножницами поглаживали область кожного покрова вокруг вибрисс (контрольная группа). В возрасте 28 дней крысят отсаживали от матери и далее содержали группами по 3–4 самца в каждой клетке. В возрасте 90 дней тестировали поведение самцов в «открытом поле». За 2 дня до теста проводили кратковременный хэндлинг животных (взятие в руки и пересадка в переноску на 5 мин) для снижения стрессогенности тестирования и повышения двигательной активности. Всего было исследовано поведение десяти контрольных и семи опытных животных.
«Открытое поле» представляло собой квадратную, равномерно освещенную арену черного цвета размером 108х108 см, огороженную стенками высотой
Для тестирования использовали видеотрекинг. Поведение животного снимали видеокамерой, установленной на высоте
Число стоек и актов груминга подсчитывали суммарно за 10 мин тестирования посредством визуального наблюдения за поведением животного на мониторе.
На первом этапе сбора данных с помощью программы «Easy Track» вычисляли следующие показатели двигательной активности: длину пройденного пути (ДП), среднюю (Vср, усредненное значение за период тестирования) и максимальную скорости движения животного (Vmax, наибольшее значение за период тестирования). Данные показатели оценивали за 10 мин теста (исходные значения и нормированные по каждому помету), а также отдельно за первую (t1=0–5 мин.) и вторую (t2=5–10 мин.) половину тестирования, для всего «открытого поля» и отдельно для центральной зоны. Для нормирования использовали следующую формулу: нормированное значение = (исходное значение – среднее значение)/√ D. Для определения типичных значений показателей поведения высчитывали средние и медианы. В качестве меры вариабельности использовали ошибку среднего (ОС) и дисперсию (D) – величины, связанные соотношением D=ОС2*N, где N – размер выборки (Zar, 2010).
Для дальнейшего анализа использовали программу MatLab (версия R2012b). Данные о перемещении крысы в «открытом поле» (зависимость координат от времени) x(t) и y(t) обрабатывали с помощью фильтра Савицкого–Голея (Savitsky–Golay), обеспечивающего оптимальное сглаживание в смысле наименьших квадратов полиномами второго порядка на окне в 25 точек. Сглаживание не искажало полученные в эксперименте траектории, что проиллюстрировано на рис. 1Б, где сглаженные траектории практически неотличимы от экспериментально полученных траекторий (рис. 1Б, пунктир).
Затем данные дифференцировали по времени для нахождения абсолютных значений скорости, v:
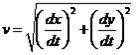
и ускорения, a:
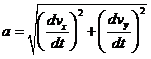 .
.
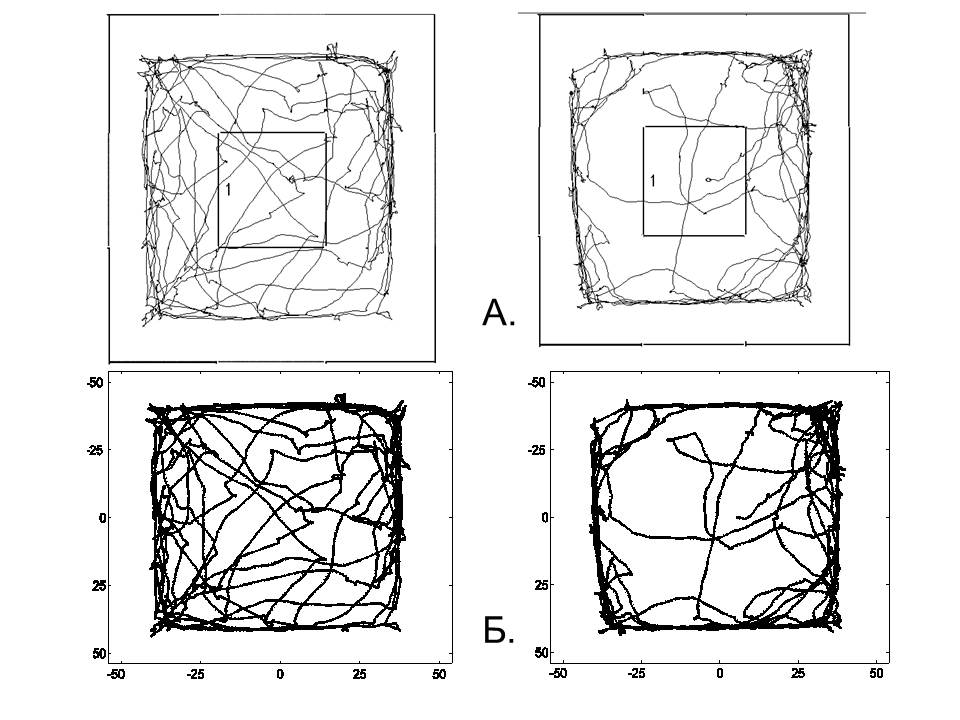
Рис. 1. Примеры траекторий движения крыс в «открытом поле» за 10 мин тестирования. На рис. А и Б слева приведены траектории контрольного животного, справа – опытного. А – траектории, построенные программой «Easy Track». Границы внешних квадратов соответствуют стенкам «открытого поля». Цифрой 1 обозначена центральная зона. Б – траектории, аппроксимированные в программе MatLab с помощью фильтра Савицкого–Голея (сплошная линия) и наложенные на оригинальные (пунктир). Видно, что траектории едва различимы. По вертикали и горизонтали указаны координаты «открытого поля»
В дополнение к названным параметрам вычисляли расстояние до ближайшей стенки, dw:
![]() ,
,
где lw=-
Для оценки вероятности обнаружения животных, обладающих определенными кинематическими характеристиками, осуществляли построение кумулятивной функции частоты встречаемости. При построении этой функции для величин v, a, dw применяли оценку плотности вероятности Каплана–Мейера (Kaplan–Meier).
При обработке данных мы использовали непараметрические малочувствительные к распределению статистические тесты Манна–Уитни (в тексте этот тест обозначен как U), сравнения медиан Муда (Mood, 1950) (в тексте обозначен как M) и сравнения дисперсий Брауна–Форсайта (Brown–Forsythe, в тексте обозначен как BF) (Brown, Forsythe, 1974). Следует отметить, что классический тест Манна–Уитни можно использовать и для оценки медиан, однако тест Муда накладывает меньше ограничений на распределения, и этому тесту стоит отдавать предпочтение при оценке достоверности различий медиан.
Статистический анализ проводили в программах STATISTICA (версия 8.0) и MatLab (версия R2012b).
Результаты
Сравнение исходных значений стандартных показателей двигательной активности (ДП, Vср и Vmax) за 10 мин теста не выявило статистически значимых различий между контрольными и опытными животными (табл. 1). При анализе данных, нормированных по пометам, было обнаружено статистически значимое уменьшение числа стоек и ДП в центральной зоне «открытого поля» у опытных крыс (табл. 1). Также выявлена тенденция к меньшей ДП во всем «открытом поле» у опытных животных по сравнению с животными контрольной группы (тест U, p=0,06).
Таблица 1
Стандартные показатели поведения контрольных и опытных крыс в тесте «открытое поле»
|
Показатель поведения |
Исходные данные |
Данные, нормированные по пометам | ||
|
Контроль N=10 |
Опыт N=7 |
Контроль N=10 |
Опыт N=7 | |
|
Длина пути общая (см) |
3109 ± 307 |
2876 ± 154 |
0,45 ± 0,30 |
-0,47 ± 0,17 |
|
Длина пути в центральной зоне (см) |
144 ± 38 |
107 ± 22 |
0,52 ± 0,29 |
-0,59 ± 0,13 |
|
Средняя скорость движения (см/c) |
5,2 ± 0,5 |
4,9 ± 0,2 |
0,44 ± 0,31 |
-0,45 ± 0,16 |
|
Максимальная скорость движения (см/с) |
47,7 ± 4,4 |
42,1 ± 1,3 |
0,11 ± 0,24 |
-0,005 ± 0,40 |
|
Число стоек |
16,2 ± 2,1 |
12,3 ± 1,9 |
0,51 ± 0,18 |
-0,53 ± 0,32 |
|
Число актов груминга |
5,4 ± 1,1 |
8,1 ± 1,1 |
-0,30 ± 0,22 |
0,47± 0,37 |
Примеч.: Жирным шрифтом выделены статистически значимо различающиеся значения для данных групп (тест U, p<0,05).
Второй этап анализа состоял в сравнении показателей активности контрольных и опытных крыс в периоды t1 и t2, поскольку во втором периоде теста двигательная активность отражает преимущественно исследовательское поведение (Маркель и др., 1988). Сначала мы проанализировали динамику двигательной активности для каждого животного. Обнаружено, что в период t2 по сравнению с t1 у 30% контрольных крыс была больше ДП во всем «открытом поле», у 60% – больше ДП в центральной зоне, у остальных контрольных животных эти показатели уменьшались. У каждого опытного животного ДП как во всем «открытом поле», так и в центральной зоне, за период t2 была меньше, чем за период t1.
Затем мы оценили изменение показателей активности животных от периода t1 к t2 по средним значениям, медианам и дисперсиям. У контрольных крыс показатели поведения в периоды t1 и t2 не различались (табл. 2, 3). У опытных крыс в период t2 значения ДП и Vcp во всем «открытом поле», а также Vmax в центральной зоне были ниже, чем в период t1 (табл. 2). Статистическая значимость различий подтверждена путем сравнения медиан (тест М, p<0,05) и U-тестом (p<0,05, табл. 3). Для остальных показателей поведения опытных крыс в периоды t1 и t2 статистически значимые различия не были выявлены (табл. 3).
Таблица 2
Кинематические характеристики двигательной активности контрольных и опытных крыс в «открытом поле» за периоды t1 и t2
|
Измере ния |
Показатель поведения |
Контроль, N=10 |
Опыт, N=7 | ||||||
|
1–5 мин |
6–10 мин |
1–5 мин |
6–10 мин | ||||||
|
СР ± ОС |
М |
СР ± ОС |
М |
СР. ± ОС |
М |
СР ± ОС |
М | ||
|
Для всего «отк-рыто-го поля» |
Длина пути (см) |
1689 ± 211 |
1682 |
1188 ± 176 |
1280 |
1699 ± 95 |
1692 |
1083 ± 109 |
1078 |
|
Средняя скорость движения (см/c) |
5,64 ± 0,70 |
5,62 |
3,98 ± 0,60 |
4,27 |
5.69 ± 0,32 |
5,65 |
3,62 ± 0,36 |
3,60 | |
|
Макси-мальная скорость движения (см/с) |
38,71 ± 3,72 |
38,0 |
36,14 ± 4,19 |
36,06 |
45,80 ± 4,11 |
45,94 |
35,94 ± 1,53
|
36,19 | |
|
В цент-раль-ной зоне |
Длина пути (см) |
42 ± 14 |
18 |
80 ± 22 |
69 |
57 ± 12 |
61 |
33 ± 11 |
44 |
|
Средняя скорость движения (см/c) |
9,88 ± 2,06 |
9,96 |
7,64 ± 2,40 |
6,14 |
9,83 ± 1,59 |
10,20 |
4,51 ± 1,51 |
5,54 | |
|
Максима-льная скорость движения (см/с) |
22,80 ± 3,48 |
21,0 |
25,50 ± 5,60 |
23,71 |
36,94 ± 6,80 |
34,97 |
14,80 ± 4,96 |
19,25 | |
Примеч.: представлены значения средних величин (СР), ошибки среднего (ОС), медианы (М); жирным шрифтом выделены значения ОС, отражающие статистически значимые различия дисперсий у контрольных и опытных крыс в соответствующем периоде тестирования (тест BF, p<0,05).
Таблица 3
Оценка статистических различий между кинематическими характеристиками двигательной активности контрольных и опытных крыс в «открытом поле» для периодов t1 и t2
|
Измерения |
Пока-затель Поведения |
Сравнение показателей поведения для периодов t1 и t2 |
Сравнение показателей поведения у контрольных (N1=10) и опытных (N2=7) крыс |
| |||||||||||
|
У контрольных N1=N2=10 |
У опытных N1=N2=7 |
Для t1 |
Для t2 | ||||||||||||
|
U |
M |
BF |
U |
M |
BF |
U |
M |
BF |
U |
M |
BF | ||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 | ||
|
Для всего «открытого поля» |
Длина пути (cм) |
0,11 |
0,37 |
0,40 |
0,01 |
0,02 |
0,35 |
1,0 |
1,0 |
0,01 |
0,52 |
0,30 |
0,18 | ||
|
Ср. скор. (cм/c) |
0,13 |
0,37 |
0,45 |
0,01 |
0,02 |
0,38 |
1,0 |
1,0 |
0,01 |
0,52 |
0,30 |
0,16 | |||
|
Макс. скор. (cм/c) |
0,76 |
1,0 |
0,49 |
0,054 |
0,25 |
0,053 |
0,23 |
0,30 |
0,99 |
1,0 |
1,0 |
0,03 | |||
|
В центральной зоне |
Длина пути (см) |
0,36 |
0,37 |
0,12 |
0,15 |
0,25 |
0,58 |
0,19 |
0,19 |
0,64 |
0,19 |
0,30 |
0,02 | ||
|
Ср. скор. (cм/c) |
0,33 |
0,32 |
0,97 |
0,11 |
0,25 |
0,56 |
1,0 |
1,0 |
0,31 |
0,59 |
1,0 |
0,40 | |||
|
Макс. скор. (cм/c) |
0,43 |
0,37 |
0,35 |
0,04 |
0,02 |
0,54 |
0,13 |
0,30 |
0,34 |
0,10 |
0,04 |
0,55 | |||
Примеч.: Ср. скор. – средняя скорость движения; Макс. скор. – максимальная скорость движения; представлены значения р для тестов U, M, BF; жирным шрифтом выделены значения p<0,05.
Далее мы сравнили контрольных и опытных животных по двигательной активности в каждый из периодов. Статистически значимых различий медиан для большинства показателей поведения в обоих периодах теста не было обнаружено (табл. 2, 3). Единственным значимым различием (табл. 3) было меньшее значение медианы Vmax в центральной зоне в период t2 у опытных животных по сравнению с животными контрольной группы (19,25 и 23,71 соответственно).
Статистически значимые различия между показателями двигательной активности контрольных и опытных животных были выявлены в отношении дисперсий. В период t1 у контрольных и опытных крыс различались дисперсии ДП и Vср в целом «открытом поле», в период t2 – дисперсии Vmax в целом «открытом поле» и ДП в центральной зоне (тест BF, p<0,05, табл. 3). У опытных крыс дисперсии всех этих показателей были значительно меньше, чем у животных контрольной группы (табл. 2). Дисперсии остальных показателей поведения не различались у контрольных и опытных животных (табл. 3).
Для детального описания кинематики движения крыс в «открытом поле» были построены кумулятивные кривые частоты встречаемости событий, т.е. оценки вероятности наблюдений определенных значений расстояния от животного до стенки (dw), скорости движения (v) и ускорения (a) (рис 2). На графиках для точки (x0,P0), ордината P0 показывает долю наблюдений, в которых показатель движения (т.е. положение, скорость или ускорение, рис. 2А, Б, В) не превышает соответствующего значения x0 (Zar, 2010).
Для всех показателей кривые, построенные по данным активности контрольных и опытных животных, имели сходную форму (рис. 2). Однако кривые, построенные на основе характеристик двигательной активности опытных крыс, располагались ближе друг к другу, узким пучком внутри более широкого пучка кривых, построенных для контрольных крыс (рис. 2).
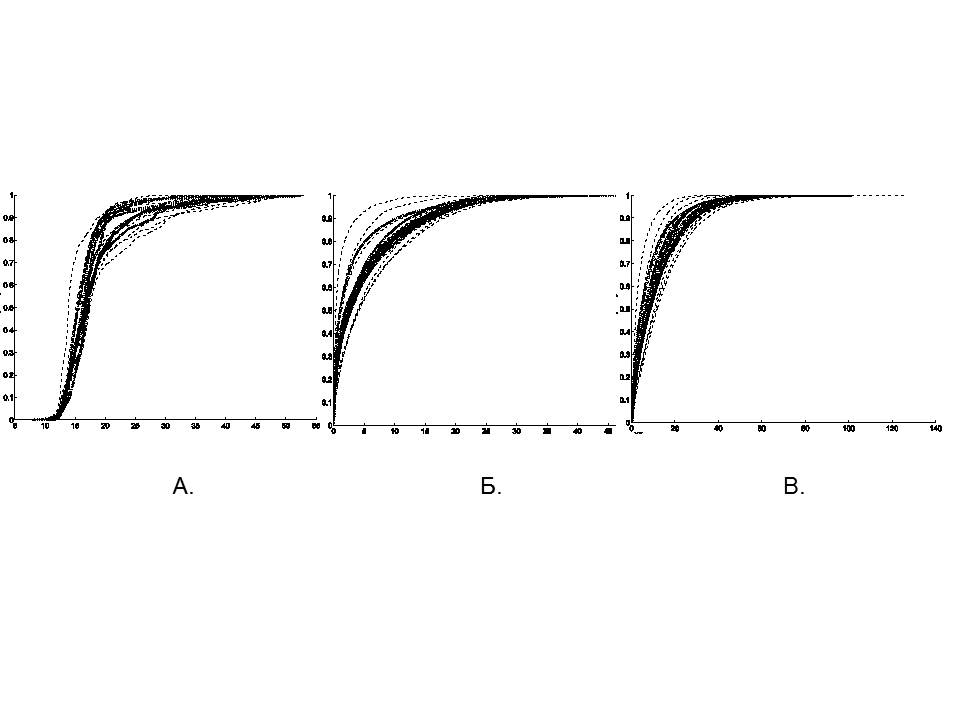
Рис. 2. Кумулятивные функции распределения расстояния до стенки (dw), скорости движения (v) и ускорения (a). Пунктирными линиями показаны кривые, построенные по данным контрольных крыс, сплошными линиями – кривые, построенные по данным опытных крыс. По горизонтали: А – расстояние от животного до стенки (см); Б – скорость движения животного (см/с); В – ускорение животного (см/с2), по вертикали на всех графиках – частота встречаемости
Для оценки статистической значимости этих наблюдений были построены распределения медиан скорости, ускорения и расстояния до стенки. Заметим, что на рис.2 медиане соответствует точка пересечения кумулятивных кривых горизонтальной линией на уровне 0.5. Оказалось, что значения медиан этих распределений у контрольных и опытных крыс близки (рис. 3), в то время как величина дисперсий для контрольной группы больше, чем для опытной.
Статистический анализ подтвердил, что различия медиан не значимы (тест М, N1=10, N2=7, p>0,05: для скорости движения p=0,58, для ускорения p=0,58, для расстояния до стенки p=0,44). В то же время различия дисперсий для скорости и ускорения оказались статистически значимыми (тест BF, N1=10, N2=7, p=0,012 и p=0,04 соответственно). При анализе показателей расстояния от животного до стенки (рис. 3) не было выявлено статистически значимых различий дисперсий (BF, N1=10, N2=7, p=0,14). Заметим, что при анализе дисперсий показателей расстояния от животного до стенки мы учитывали и значение единичного выброса (рис. 3А, кружок).
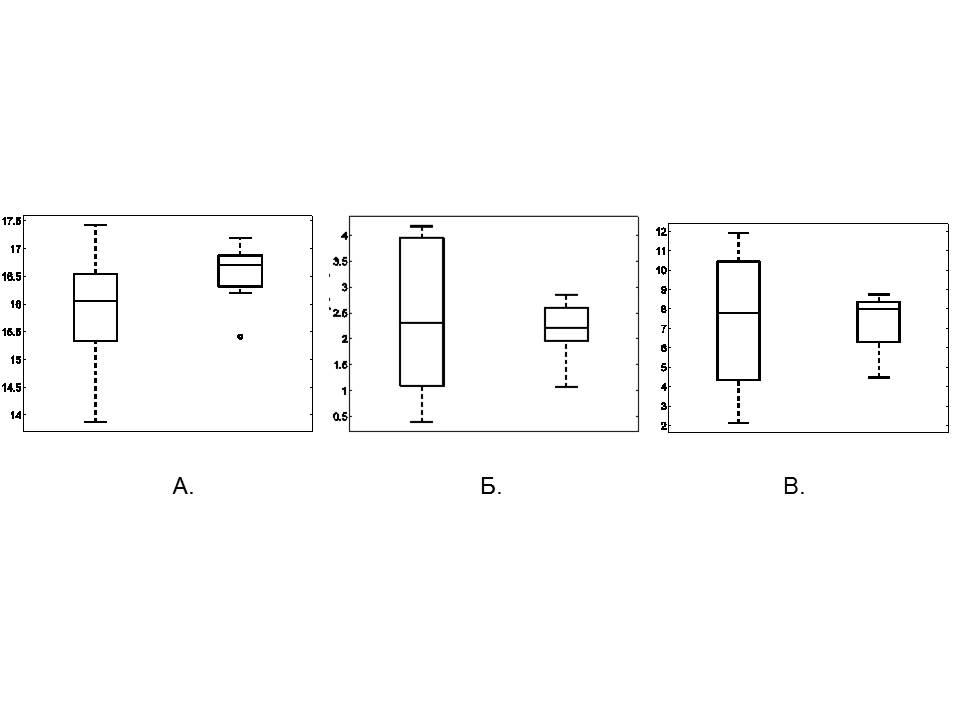
Рис. 3. Сравнение медиан и дисперсий: А – расстояния от животного до стенки (см); Б – скорости движения животного (см/с); В – ускорения движения животного (см/с2). На каждом рисунке слева – данные контрольной группы, справа – данные опытной группы. Нижняя и верхняя границы прямоугольников соответствуют 25 и 75 процентилям. Линия внутри прямоугольника – 50 процентиль (медиана). Линии, выходящие за прямоугольник, указывают разброс значений соответствующей величины. Кружком на рис. А указан единичный выброс
Обсуждение
В настоящей работе мы проанализировали влияние снижения видоспецифического сенсорного притока от вибрисс с 9-го по 20-й дни жизни на кинематические параметры двигательной активности взрослых крыс в «открытом поле». Отправной точкой исследования было предположение, что отдаленный эффект ограничения видоспецифической афферентации наиболее вероятно будет выявляться в целенаправленном поведении взрослых животных в условиях «свободного поведения» в среде, требующей проявления активности для адаптации к новым условиям (Крылов, Александров, 2011).
Полученные данные указывают на то, что выстригание вибрисс у крысят с 9-го по 20-й день жизни не вызывает существенных изменений общего уровня их двигательной активности в трехмесячном возрасте по показателям скорости движения (средней и максимальной) и длины пути.
В то же время нами был обнаружен ряд различий между контрольными и вибриссэктомированными крысами в динамике и организации двигательной активности. Это было характерно как для первого, так и для второго 5-минутных периодов тестирования. Однако в связи с тем, что поведение крыс в течение первых 5 мин в «открытом поле» в равной степени отражает как тревожность в незнакомой обстановке, так и исследовательскую активность (Маркель и др., 1988), изменение вариабельности параметров перемещений в этот период может быть проявлением нарушений как оборонительного, так и исследовательского видов поведения и/или взаимодействия между ними. Результаты последующего 5-минутного тестирования наиболее специфично отражают исследовательские реакции. Полученные данные свидетельствуют о том, что экспериментальных животных отличает меньшая двигательная активность по всем исследованным показателям: длина пути, средняя скорость движения во всем «открытом поле» и максимальная скорость движения в центральной зоне.
Уменьшение пройденного пути у опытных крыс, особенно в центральной (потенциально более опасной) зоне, свидетельствует о более низком уровне исследовательской активности. В пользу такого вывода свидетельствуют и полученные нами данные об уменьшении числа стоек у вибриссэктомированных крыс. Следует подчеркнуть, что обнаруженное явление характерно для ограничения сенсорного притока с 9-го по 20-й день жизни, когда вибриссэктомия нарушает синхронность формирования ранних поведенческих реакций. Удаление вибрисс с момента рождения приводит не к исчезновению, а к реорганизации корреляционных связей. Прозревание у таких депривированных крысят начинается раньше, и его темпы коррелируют с развитием большинства ранних поведенческих реакций (Шишелова, Раевский, 2009), что отражает адаптивные компенсаторные перестройки нейронных сетей мозга. Следовательно, обнаруженное Ли и соавт. увеличение пройденного пути и числа стоек у одномесячных крыс, подвергавшихся вибриссэктомии с 1-го по 4-й день жизни, имеет не случайный характер (Lee at al., 2009).
Наряду с этим, полученные результаты свидетельствуют о меньшей вариативности двигательной активности экспериментальных животных по сравнению с контрольными крысами: как по дисперсии исследованных показателей, так и по группировке кривых кумулятивных функций их распределения. Метод построения кумулятивных функций оказался эффективен для поставленной нами задачи и показал, что, несмотря на сходство средних динамических характеристик двигательной активности контрольных и опытных животных, последние ведут себя более стереотипно. В основе данного явления может лежать сужение границ проявления исследовательского поведения, подразумевающего разнообразие движений (скоростей) и резкости движений (ускорений) у депривированных крыс по сравнению с контрольными.
Кроме этого, временная динамика пройденного пути у всех опытных крыс была одинаковой и выражалась в уменьшении длины пути, пройденного за второй период, по сравнению с первым, а в группе контрольных крыс у части животных наблюдали уменьшение и у части – увеличение пройденного пути. Такого рода закономерность также позволяет говорить о меньшем разнообразии проявлений исследовательского поведения у депривированных животных.
Известно, что временная сенсорная депривация обусловливает ограничение учета деталей среды при организации поведения (Alexandrov, 2008). Вполне возможно, что наблюдаемое нами уменьшение разнообразия двигательной активности у животных, перенесших временную вибриссэктомию в раннем онтогенезе, происходит за счет менее структурированного анализа окружающей среды.
В целом результаты исследования свидетельствуют о том, что ограничение видоспецифического сенсорного притока в период с 9-го по 20-й день постнатального онтогенеза обусловливает более стереотипное поведение взрослых животных в условиях новой обстановки.
Вывод
Ограничение видоспецифической соматосенсорной афферентации у крыс с 9-го по 20-й день постнатального онтогенеза вызывает уменьшение выраженности и разнообразия проявлений исследовательского поведения в зрелом возрасте.
Литература
- Крылов А.К., Александров Ю.И. Методы экспериментального исследования в парадигмах активности и реактивности // Современная экспериментальная психология: в 2 т. / Под ред. В. А. Барабанщикова. М.: Изд-во «Институт психологии РАН», 2011. Т. 1. С. 463–478.
- Маркель А.Л., Галактионов Ю.К., Ефимов В.М. Факторный анализ поведения крыс в тесте открытого поля // Журнал высшей нервной деятельности. 1988. Т. 38. № 5. С. 855–863.
- Сварник О.Е., Булава А.И., Фадеева Т. А., Александров Ю.И. Закономерности реорганизации опыта, приобретенного при одно- и многоэтапном обучении // Экспериментальная психология. 2011. Т. 4. № 2. С. 5–13.
- Созинов А. А., Крылов А. К., Александров Ю.И. Эффект интерференции в изучении психологических структур // Экспериментальная психология. 2013. Т. 6. № 1. С. 5–47.
- Шишелова А.Ю., Раевский В.В. Влияние вибриссэктомии в раннем постнатальном онтогенезе у крысят на развитие поведения // Журнал высшей нервной деятельности. 2009. Т. 59. № 3. С. 326–334.
- Aleхandrov Yu.I. How we fragment the world: the view from inside versus the view from outside // Social Science Information. Spec. issue: Cognitive technologies. 2008. Vol. 47. № 3. P. 419–457. doi: 10.1177/ 0539018408092580
- Ardiel E.L., Rankin C.H. The importance of touch in development // Pediatr. Child Health. 2010. Vol. 15. № 3. P. 153–156.
- Bolles R.C., Woods P.J. The ontogeny of behaviour in the albino rat // Animal Behavior. 1964. Vol. 12I. № 4. P. 427–441.
- Brown, M.B., Forsythe A.B. Robust Tests for Equality of Variances // Journal of the American Statistical Association. 1974. Vol. 69. P. 364–367.
- Curley J.P., Davidson S., Bateson P., Champagne F.A. Social enrichment during postnatal development induces transgenerational effects on emotional and reproductive behavior in mice // Frontiers in behavioral neuroscience. 2009. Vol. 3. P. 1–14. doi: 10.3389/neuro.08.025.2009.
- Curley J.P., Jensen C.L., Mashoodh R.,
- denenberg v.h. Evolution proposes and ontogeny disposes // Brain and Language. 2000. Vol. 73. P. 274–296.
- Grant R.A., Mitchinson B.,
- Grant R.A., Sperber A.L., Prescott T.J. The role of orienting in vibrissal touch sensing // Frontiers in behavioral neuroscience. 2012. Vol. 6. P. 1–12. doi: 10.3389/fnbeh.2012.00039.
- Erzurumlu R.S. Critical period for the whisker-barrel system // Exp Neurol. 2010. Vol. 222. № 1. P. 10–12. doi: 10.1016/j.expneurol.2009.12.025.
- Landers M. S., Sullivan R. M. The Development and Neurobiology of Infant Attachment and Fear // Dev Neurosci. 2012. Vol. 34. № 2, 3. P. 101–114.
- Lee L.J., Chen W.J., Chuang Y.W., Wang Y.C. Neonatal whisker trimming causes long-lasting changes in structure and function of the somatosensory system // Exp Neurol. 2009. Vol. 219. P. 524–532. doi: 10.1016/j.expneurol.2009.07.012.
- Miyazaki T., Takase K., Nakajima W., Tada H., Ohya D., Sano A., Goto T., Hirase H., Malinow R., Takahashi T. Disrupted cortical function underlies behavior dysfunction due to social isolation // J Clin Invest. 2012. Vol. 122. № 7. P. 2690–2701. doi: 10.1172/JCI63060.
- Mood A.M. Introduction to the Theory of Statistics // McGraw-Hill Book Co, N. Y, 1950. 433 p.
- Pryce C.R., Feldon J. Long-term neurobehavioural impact of the postnatal environment in rats: manipulations, effects and mediating mechanisms // Neurosci Biobehav Rev. 2003. Vol. 27. № 1, 2. P. 57–71.
- Rodel H. G., Meyer S. Early development influences ontogeny of personality types in young laboratory rats // Developmental Psychobiology. 2011. Vol. 53. P. 601–613. doi: 10.1002/dev.20522.
- Shoykhet M., Simons D.J. Development of thalamocortical response transformations in the rat whisker-barrel system // J Neurophysiol. 2008. Vol. 99. P. 356–366.
- Zar J.H. Biostatistical Analysis. 5th Edition. N.J. Person PrenticeHall,
Информация об авторах
Метрики
Просмотров
Всего: 3482
В прошлом месяце: 8
В текущем месяце: 18
Скачиваний
Всего: 897
В прошлом месяце: 3
В текущем месяце: 1