Экспериментальная психология
2015. Том 8. № 1. С. 85–93
ISSN: 2072-7593 / 2311-7036 (online)
Методика изучения схемы тела у змей*
Аннотация
Общая информация
* Работа поддержана грантом РГНФ № 14-06-00669, тип проекта – а.
Ключевые слова: методика для изучения схемы тела, экспериментальная установка, змеи, пресмыкающиеся, беспозвоночные
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
Для цитаты: Хватов И.А., Соколов А.Ю., Харитонов А.Н. Методика изучения схемы тела у змей // Экспериментальная психология. 2015. Том 8. № 1. С. 85–93.
Полный текст
При ориентации в окружающем пространстве для осуществления локомоции и манипуляции животным необходимо учитывать физические характеристики собственного тела – границы, объем, массу – и соотносить их с физическими характеристиками внешних объектов. Иначе говоря, животным необходимо «принимать себя в расчет» (Столин, 1983). Человек и другие приматы осуществляют этот процесс преимущественно с опорой на зрение – для нас зрительная система является ведущей при построении когнитивной репрезентации внешнего мира и самих себя. Поэтому одним из существенных критериев сознания считается способность узнавать собственное отражение в зеркале. Этот феномен в настоящий момент, начиная с классических экспериментов Г. Гэллапа (Gallup 1970), активно исследуется и на человеческих детях, и на ряде других млекопитающих. На сегодняшний день установлен факт наличия способности к самоузнаванию у понгид, макак-резусов, слонов, дельфинов, касаток и сорок (подробнее см.: Хватов, 2013). Следует отметить, что данный метод подвергается существенной критике: во-первых, его трудно применять по отношению к животным, у которых зрительная модальность не является ведущей, (например, к большинству млекопитающих; см.: Bekoff, 2001), во-вторых, как животные, так и человек могут и не проявлять интереса к отметкам, наносимым на тело, что используется в таких исследованиях как один из основных экспериментальных приемов (Asendorpf et al., 1996), в-третьих, дискуссионным остается вопрос о том, действительно ли самоузнавание в зеркале свидетельствует о самосознании (там же).
В рамках близкого направления исследований изучается «схема тела» человека и других животных (Herman et al. 2001; Maravita, Iriki, 2004). Схема тела – это совокупность двигательных навыков и способностей, позволяющих осуществлять различные движения, а также представление о физических характеристиках собственного тела (его границах, весе, взаиморасположении отдельных членов). В отличие от образа тела, схема тела является неосознаваемым феноменом (Gallagher, Cole, 1995). Существует точка зрения, что схема тела («принятие себя в расчет») является филогенетически наиболее ранней ступенью развития всех прочих представлений о себе, включая самосознание (Столин, 1983). Ранее было показано, что животные способны включать в схему собственного тела внешние объекты (Maravita, Iriki, 2004).
На сегодняшний день в науке отсутствует отработанная методика, позволяющая ставить поведенческие эксперименты на пресмыкающихся с целью изучения особенностей их схемы тела. Однако за последние годы коллективом авторов данной статьи была разработана оригинальная методика, нацеленная на исследование специфики схемы тела у беспозвоночных. Ниже будет приведено краткое описание данной методики.
В ходе экспериментов изучалось влияние изменения физических параметров тела животных на их поведение в различных экспериментальных установках. На начальном этапе эксперимента животное помещалось в экспериментальную установку (лабиринт или проблемную клетку) с целью формирования у него привыкания к данным условиям и осуществлению в них своей жизнедеятельности – в первую очередь кормления. Далее границы тела животного увеличивались экспериментатором с помощью крепления на него различных объектов таким образом, что они препятствовали осуществлению животным ранее сформированных двигательных навыков – проникновению в отверстия в перегородках, отделяющих один отсек лабиринта или проблемной клетки от другого. На других этапах эксперимента, наоборот, изменялись физические параметры среды (размеры отверстий в лабиринтах и проблемных клетках) при неизменности границ тела животного. Анализ данных о локомоторной активности животных, полученных в ходе этих экспериментов, позволил определить, способны ли животные учитывать физические параметры собственного тела при осуществлении поведения, а также способны ли они научаться менять свое поведение при изменении этих параметров и экстраполировать приобретенный опыт в новые ситуации. С помощью этой методики были получены данные о специфике психического отражения параметров собственного тела у тараканов, сверчков и моллюсков (Хватов, 2010; 2011; Хватов, Харитонов, 2012, 2013; Хватов, Харитонов, Соколов, 2013).
На основе данной методики была также разработана и апробирована на крысах методика для исследования схемы тела у мелких млекопитающих (Хватов и др., 2014).
В настоящей работе мы предлагаем описание и апробацию модифицированного варианта методики и новой экспериментальной установки, ориентированной на исследование схемы тела змей, которая была разработана с учетом опыта наших исследований.
Метод
В качестве объекта исследования предполагается использование змей различных видов. Змеи являются весьма удобным объектом для экспериментов описанного выше типа, поскольку проникновение в различные типы отверстий при измененных (увеличенных) границах тела является для этих животных экологичной задачей: при проглатывании добычи (например, мыши или крысы) размер их тела существенно увеличивается, что избавляет экспериментатора от необходимости закреплять на теле животного какие-либо дополнительные предметы.
Ранее змеи не выступали объектами такого рода исследований. В настоящее время основная часть работ, проводимых на этих пресмыкающихся, посвящена исследованию особенностей их биомеханики (Jayne, Riley, 2007; Marvi, Hu, 2012; Xiao, Chen, 2013). В поведенческих экспериментах со змеями традиционно исследуется их поведение избегания в лабиринте с последующим проникновением в комфортное помещение (Kellogg, Pomeroy, 1936; Crawford, Bartlett, 1966; Fuenzalida, Ulrich, 1975). Используя эту особенность поведения змей, мы создали для наших исследований экспериментальную установку, в которой животному необходимо учитывать границы собственного тела при проникновении в различные отверстия.

Рис. 1.Схема экспериментальной установки: внизу отсек №1 (светлый), вверху отсек №2 (темный); номерами обозначены отверстия (см. в тексте)
Оборудование. Экспериментальная установка представляет собой стеклянный ящик с двумя отсеками (рис. 1).
-
Отсек №1 – пусковая камера 310x390 мм. Пол покрыт шпатовой и мраморной окатанной галькой белого цвета (для удобства различения змеи на субстрате при видеосъемке).
-
Отсек №2 – «влажная» камера (укрытие) 250x390 мм. В качестве субстрата использовалась сосновая кора-мульча, сфагнум, крошка кокосовой коры. Субстрат увлажнялся исходно и затем дополнительно каждые 2 дня. В этом отсеке располагалась поилка. Стены и потолок отсека с внешней стороны были окрашены черной светонепроницаемой краской, благодаря чему в нем постоянно поддерживался низкий уровень освещенности.
Камеры разделены между собой стеклянной перегородкой, также окрашенной в черный цвет. В перегородке имеются три круглых отверстия диаметром
В эксперименте, проводившемся для апробации методики и установки, использовались отверстия трех диаметров:
-
большое отверстие (D=70 мм, без вставки) – в данное отверстие змея могла проникнуть без затруднений даже после проглатывания пищи;
-
среднее отверстие (вставка с отверстием D=14 мм) – в данное отверстие змея проникала без затруднений за исключением ситуации, когда ее тело было увеличено после заглатывания пищи;
-
малое отверстие (вставка с отверстием D=8 мм) – в данное отверстие змея проникнуть не могла.
Расположение отверстий в лабиринте: №1 – большое, №2 – среднее, №3 – малое.
Апробация
Животные: 3 самца и 1 самка королевской змеи Lampropeltis triangulum campbelli. Для данного вида змей является экологичным роющее поведение, они предпочитают находиться в темном и влажном окружении.
Фиксируемые показатели: время решения экспериментальной задачи (от момента помещения животного в отсек №1 до момента, когда его тело целиком покинет данный отсек через одно из отверстий), количество проникновений/попыток проникновения в каждое из трех отверстий.
Цель исследования: установить, способны ли змеи формировать навык проникновения в отсек № 2 («влажную» камеру), о чем должно было свидетельствовать сокращение времени решения задачи, а также сокращение попыток проникновения в отверстия различных диаметров.
Процедура эксперимента. Перед началом эксперимента змея в течение 7 дней содержалась в экспериментальной установке, где все три отверстия имели диаметр

Рис. 2. Змея, делающая попытку проникновения во «влажную» камеру экспериментальной установки
Результаты. У всех испытуемых от 1-й к 20-й пробе наблюдалось снижение времени решения задачи: на 1-й пробе у экспериментальной выборки среднее время решения составило 327 с (SD=38,60), на 20-й пробе – 12,5 с (SD=3,41) (рис. 3). Также у всех змей к 20-й пробе снижалось количество попыток проникновения в различные отверстия: за первые 5 проб общее количество попыток проникновения в отверстие №1 – 20, в отверстие №2 – 43, в отверстие №3 – 34. За последние 5 проб общее количество попыток проникновения в отверстие №1 – 20, в отверстие №2 – 3, в отверстие №3 – 0. Распределения достоверно отличаются друг от друга (χ2=37,35; df=2; p<0,01). Соответственно, к концу данного этапа змеи стали достоверно чаще совершать попытки проникновения в отверстие №1 и переставали пробовать проникнуть в отверстие № 3.
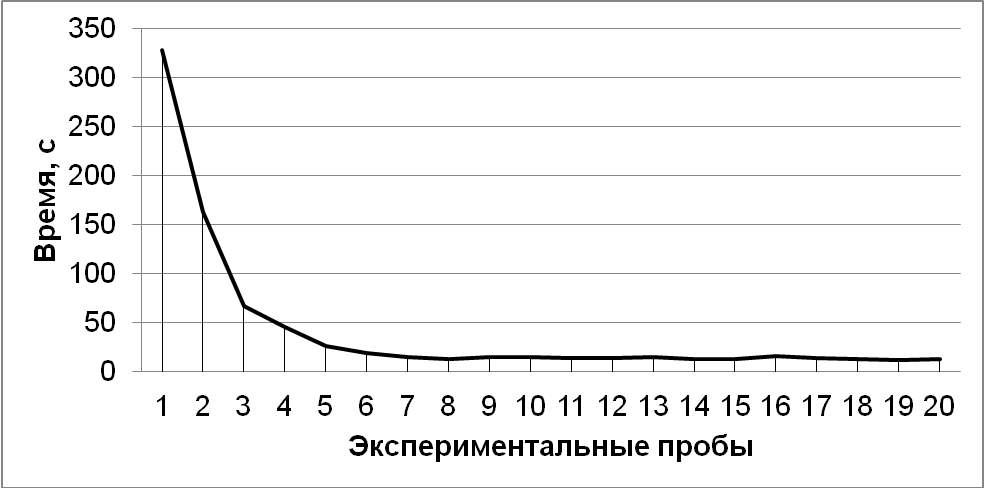
Рис. 3. Среднее время, затраченное на решение экспериментальной задачи
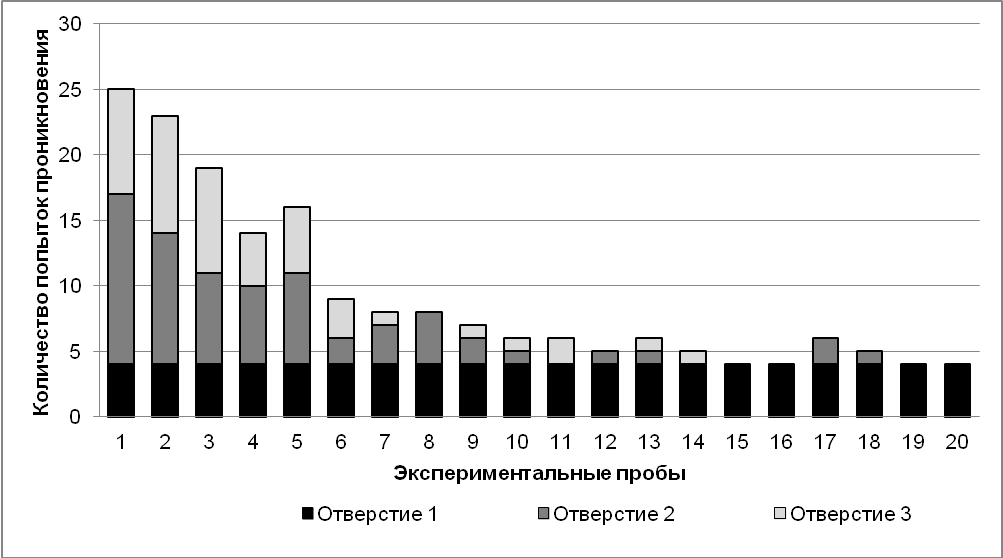
Рис. 4. Общее количество попыток проникновения в различные отверстия
Обсуждение
Полученные данные свидетельствуют о том, что у змей формируется навык проникновения кратчайшим путем в отсек №2 через отверстие №1. Соответственно, данная экспериментальная установка может быть использована для изучения особенностей научения у змей.
Для исследования схемы тела у животных предполагается ввести дополнительную экспериментальную переменную – варьирование границ тела змей путем использования экспериментальных животных после кормления, что, как это было отмечено выше, избавляет от необходимости закреплять на теле животного какие-либо инородные объекты.
Данные, полученные в ходе предыдущих исследований, проводившихся на беспозвоночных (Хватов, 2010; 2011; Хватов, Харитонов, 2012, 2013, Хватов, Харитонов, Соколов, 2013), позволяют заключить, что у ряда видов этой таксономической группы отсутствует схема тела в качестве целостной самостоятельной когнитивной структуры, данная схема складывается у них ситуативно в качестве элемента определенной перцептивной системы (целостного образа ситуации), формирующегося в процессе взаимодействия субъекта с конкретными условиями внешней среды. Если целостный образ ситуации у этих животных являлся перцептивным – в частности, в его структуре интегрируется визуальная и кинестетическая информация, – то отражение собственных характеристик как компонентов этого образа остается сенсорным, поскольку формируется на основе одномодальной информации, преобладающей по силе и поступающей из определенного сегмента тела.
На основе данных об основных тенденциях эволюции психики позвоночных и, в частности, млекопитающих (Филиппова, 2012; Хватов, 2012а; 2012b) можно выдвинуть гипотезу о том, что у позвоночных – возможно, уже у хладнокровных (пресмыкающихся) – возникает перцептивная схема тела, формирующаяся на основе интеграции сенсорных сигналов различных модальностей, поступающих от отдельных частей тела. Это позволяет животному антиципировать возможные последствия взаимодействия собственного тела с объектами в окружающем пространстве. Возможно, такая особенность в ходе прогрессивной эволюции привела к формированию обобщенного психического образа собственного тела у млекопитающих и птиц, и именно это качественно отличает психику позвоночных от психики беспозвоночных, схема тела которых представляет собой совокупность сенсорных сигналов, не связанных (или очень слабо связанных) между собой.
На наш взгляд, предложенная в настоящей статье методика может быть использована в ходе сравнительно-психологических экспериментов, нацеленных, в частности, на проверку вышеизложенной гипотезы.
Литература
- Столин В.В. Самосознание личности. М.: Издательство Московского Университета, 1983. 284 с.
- Филиппова Г.Г. Зоопсихология и сравнительная психология: учеб. пособие для студентов вузов. 6-е изд., перераб. М.: Академия, 2012. 543 с.
- Хватов И.А. Особенности самоотражения у животных на разных стадиях филогенеза: дисс. канд. психол. наук. М., 2010.
- Хватов И.А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011. Т. 4. №
- Хватов И.А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 1 [Электронный ресурс] // Психологические исследования. 2012а. № 1 (21). С. 1. URL: http://psystudy.ru/index.php/num/2012n1-21/622-khvatov21.html (дата обращения: 22.09.2014). doi: 0421200116/0001.
- Хватов И.А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 2 [Электронный ресурс] // Психологические исследования. 2012b. № 2 (22). С. 12. URL: http://psystudy.ru/index.php/num/2012n2-22/648-khvatov22.html (дата обращения: 22.09.2014). doi: 0421200116/0024.
- Хватов И.А. Проблема интеллекта животных в контексте структурно-интегративного и дифференционно-интеграционного подходов [Электронный ресурс] // Психологические исследования. 2013. Т. 6, № 28. С. 1. URL: http://psystudy.ru/index.php/num/2013v6n28/788-khvatov28.html (дата обращения: 28.07.2014).
- Хватов И.А., Харитонов А.Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. Т. 5. №
- Хватов И.А., Харитонов А.Н. Модификация плана развертки собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica // Экспериментальная психология. 2013. Т. 6. № 2. С. 101–114.
- Хватов И.А., Харитонов А.Н., Соколов А.Ю. Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Gryllus assimilis // Экспериментальная психология. 2013. Т. 6. № 4. С. 79–95.
- Хватов И.А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Методика изучения схемы тела у мелких млекопитающих // Экспериментальная психология. 2014. Т. 7. №.3. С. 137–144.
- Asendorpf J.B., Warkentin V., Baudonniere P.M. Self-Awareness and Other-Awareness II: Mirror Self-Recognition, Social Contingency Awareness, and Synchronic Imitation // Developmental Psychology. 1996. Vol. 32. № 2. P. 313–321.
- Bekoff M. Observations of scent-marking and discriminating self from others by a domestic dog (Canis familiaris): tales of displaced yellow snow // Behavioural Processes. 2001. Vol. 55. № 2. P. 75–79.
- Crawford F.T.,
- Fuenzalida C.E., Ulrich G. Escape learning in the plains garter snake, Thamnophis radix // Bulletin of the Psychonomic Society. 1975. Vol. 6(2), P. 134–136.
- Gallagher S., Cole J. Body Schema and Body Image in a Deafferented Subject // Journal of Mind and Behavior. 1995. Vol. 16. P. 369–390.
- Herman L.M., Matus D.S., Herman E.Y.K., Ivancic M., Pack A.A. The bottlenosed dolphin’s (Tursiops truncatus) understanding of gestures as symbolic representations of its body parts // Animal Learning & Behavior. 2001. Vol. 29. № 3. P. 250–264.
- Jayne B.C., Riley M.A. Scaling of the axial morphology and gap-bridging ability of the brown tree snake, Boiga irregularis // J Exp Biol. 2007. Vol. 210. P. 1148–1160.
- Kellogg W.N., Pomeroy W.B. Maze learning in water snakes // Journal of Comparative Psychology. 1936. Vol. 21(3), P. 275–295. doi: 10.1037/h0063023.
- Maravita A., Iriki A. Tools for the body (schema) // Trends in Cognitive Sciences. 2004. Vol. 8. № 2. P. 79–86.
- Marvi H., Hu D.L. Friction enhancement in concertina locomotion of snakes // J.R. Soc. Interface. 2012. doi:10.1098/rsif.2012.0132 Published online.
- Xiao J., Chen X. Buckling morphology of an elastic beam between two parallel lateral constraints: implication for a snake crawling between walls // J.R. Soc. Interface. 2013 V. 10(85). doi: 10.1098/rsif.2013.0399.
Информация об авторах
Метрики
Просмотров
Всего: 3863
В прошлом месяце: 28
В текущем месяце: 57
Скачиваний
Всего: 790
В прошлом месяце: 3
В текущем месяце: 3