Экспериментальная психология
2016. Том 9. № 1. С. 112–130
doi:10.17759/exppsy.2016090109
ISSN: 2072-7593 / 2311-7036 (online)
Схема собственного тела у грызунов (на примере крыс Rattus norvegicus)*
Аннотация
Общая информация
* Исследование проводится при финансовой поддержке Роснауки (проект № МК-5915.2014.6)
Ключевые слова: схема тела, самоотражение, тело, грызуны, крысы, филогенез психики
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2016090109
Для цитаты: Хватов И.А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Схема собственного тела у грызунов (на примере крыс Rattus norvegicus) // Экспериментальная психология. 2016. Том 9. № 1. С. 112–130. DOI: 10.17759/exppsy.2016090109
Полный текст
СОКОЛОВ АЛО.**, Лаборатория-студия -«Живая Земля»-, Москва, Россия, e-mail: apophis-king@mail.ru
ХАРИТОНОВ А.Н.***, Институт психологии РАН; Московский городской психолого-педагогический университет, Москва, Россия, e-mail: ankhome47@list.ru
КУЛИЧЕНКОВА К.Н.****, МГУ имени М.В. Ломоносова, Москва, Россия, e-mail: koulitchenkova@gmail.com
Схема собственного тела у крыс Rattus norvegicus изучалась с помощью методики, по которой животным было необходимо учитывать увеличенные границы собственного тела. Крысы экспериментальной и контрольной групп обучались доставать приманку путем проникновения головой в отверстие определенного диаметра. Затем границы тела крыс экспериментальной группы увеличивались с помощью крепления на теменную часть их головы цилиндрического объекта, препятствовавшего проникновению в те же отверстия. Для крыс контрольной группы размер отверстия, через которое животные были обучены доставать приманку, уменьшали, исключая проникновение в него головы крысы. В последующих сериях варьировалось расположение отверстий в экспериментальной установке. Показано, что при увеличении границ тела крысы способны формировать новое научение, адекватное изменившейся ситуации, что выражалось в достоверном сокращении времени решения задачи к концу каждой серии и количества неуспешных попыток. Крысы экспериментальной группы продемонстрировали также способность переносить ранее приобретенный опыт на новые ситуации, в которых изменялось расположение отверстий в экспериментальной установке. Полученные данные свидетельствуют о том, что крысы способны модифицировать схему собственного тела при изменении его физических границ.
Введение
При ориентации в окружающем пространстве для регуляции поведения животным необходимо учитывать физические характеристики собственного тела: границы, объем, массу - и соотносить их с физическими характеристиками внешних объектов. Иначе говоря, при ориентации во внешней среде животные опираются на схему собственноого тела. Схема тела - это совокупность двигательных навыков и способностей, позволяющих осуществлять различные движения, а также представление о физических характеристиках собственного тела. В отличие от образа тела, схема тела является неосознаваемым феноменом (Gallagher, Cole, 1995). Значительная часть исследований схемы тела и возможностей ее модификации проводится на людях и других приматах (Maravita, Iriki, 2004; Johnson-Frey, 2004). Недавние исследования показали, что схема тела обладает высокой дифференцированностью и гибкостью, интегрируя в свою структуру внешние объекты, находящиеся в физическом контакте с индивидом - например, орудия, используемые им (Carlson et al. 2010; Gozli, Brown, 2011; Ritchie, Carlson, 2013; Costantini et al., 2014; Moeller et al., 2015; Garbar- inia et al., 2015). Сообразно изменению схемы тела происходит и модификация субъективного восприятия окружающей реальности - эгоцентрической системы координат (Барабанщиков, 2002; Berti, Frassinetti, 2000; Costantini et al., 2014; Giglia et al., 2015). Между тем, в научной литературе отсутствуют публикации, описывающие исследования феномена схемы тела и ее изменений у большинства видов млекопитающих, включая грызунов.
Более 10 лет одним из авторов данной статьи разрабатывается концепция самоот- ражения животных и человека (Хватов, 2014), базирующаяся на онтологическом подходе (Барабанщиков, 2002). В рамках данной концепции схема тела рассматривается как один из продуктов процесса самоотражения - процесса и результата отражения субъектом своей внутренней объективной реальности (включая физические характистики собственного тела). Самоотражение рассматривается системно в контексте взаимосвязи с отражением внешней среды в целом, а, следовательно, является одной из главных составляющих единого психического отражения индивидом акта своего co-бытия с окружающей объективной реальностью.
Авторским коллективом настоящей статьи было дано целостное описание процесса развития самоотражения в ходе филогенеза животных (Хватов, 2014), а также в рамках филогенеза отдельных крупных таксонов (Хватов и др., 2014). Также был проведен ряд экспериментальных исследований особенностей процесса самоотражения и схемы тела у отдельных видов животных (Хватов, 2011; Хватов, Харитонов, 2012; Хватов и др., 2013; 2014а; 2014b). В частности, нами были разработаны и прошли успешную апробацию оригинальные экспериментальные методики изучения особенностей схемы тела у змей (Хватов и др., 2015) и мелких млекопитающих (Хватов и др., 2014а; 2014b). В рамках настоящей статьи с помощью этой методики было осуществлено полномасштабное исследование схемы собственного тела у серых крыс {Rattus norvegicus).
Гипотеза исследования. Мы полагали, что изменения границ тела крыс в сторону ограничения/препятствования осуществлению животными ранее выученного поведения будут приводить к модификации ими схемы своего тела и перестроению прежнего поведения с учетом изменений собственного тела.
Цель исследования: изучить влияние объективных изменений границ тела крыс на особенности формирования и трансформации ими схемы собственного тела.
Методика исследования
Испытуемые животные: Крысы и мыши являются модельными объектами в поведенческих исследованиях с применением различных типов лабиринтов (Barnes, 1979; Morris, 1984). Экологически и этологически обоснованной задачей для этих животных является проникновение в отверстия. Это обстоятельство используется в конструкциях некоторых лабиринтов (Barnes, 1979). Для наших исследований мы создали экспериментальную установку, в которой животному было необходимо учитывать границы собственного тела при проникновении в различные отверстия для достижения приманки.
В эксперименте было использовано 16 крыс породы Long-Evans, самцы в возрасте от 2 до 6 мес.
Животные были разделены на 2 группы: экспериментальную (8 особей) и контрольную (8 особей).
Оборудование. Экспериментальная установка представляет собой стеклянный лабиринт квадратной формы (со стороной 720 мм) с пятью отсеками: центральным отсеком квадратной формы (со стороной 200 мм) и четырьмя отсеками в форме равнобедренных трапеций, обрамляющими его. Обрамляющие отсеки сообщались между собой аркообразными отверстиями в форме усеченного круга диаметром 100 мм, расположенными в центре боковых сторон отсеков (рис. 1). Центральный отсек сообщался с каждым из четырех обрамляющих отсеков круглыми отверстиями диаметром 80 мм, располагавшимися по центру каждой из сторон отсека на высоте 5 мм от пола. Диаметр отверстий центрального отсека можно было уменьшать с помощью дополнительных вставок.
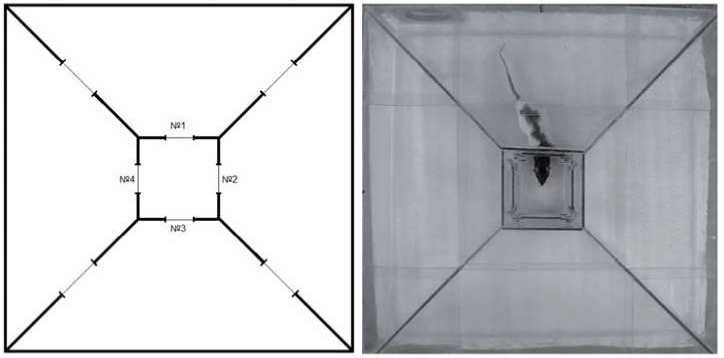
Рис 1. Слева - схема экспериментальной установки. Номерами обозначены отверстия, сообщающие внешние отсеки с центральным
В эксперименте использовались три различных диаметра отверстий центрального отсека:
• большое отверстие (D=80 мм, без вставки) - данное отверстие крыса могла проникнуть всем телом (далее - L);
• среднее отверстие (вставка с отверстием D=40 мм) — в данное отверстие могла проникнуть только голова и шея крысы (далее - М);
• малое отверстие (вставка с отверстием D=18 мм) - в данное отверстие могла проникнуть только передняя часть морды крысы (далее - S).
Процедура эксперимента. В начале каждой экспериментальной пробы крыса помещалась во внешний отсек с отверстием № 1. В центральный отсек помещалась приманка - сыр. Проба считалась завершенной после того, как крыса доставала приманку через одно из отверстий, соединяющих обрамляющие отсеки с центральным. Время пробы не ограничивалось. После каждой пробы осуществлялась очистка экспериментальной установки с применением хозяйственного мыла.
Эксперимент состоял из пяти серий для экспериментальной выборки и трех серий для контрольной выборки (табл. 1). Каждая экспериментальная серия включала в себя 20 проб. Экспериментальные серии проводились последовательно с каждым животным по две пробы ежедневно в 9 и 18 ч.
Таблица 1
|
Серии |
Группы |
|
|
Экспериментальная |
Контрольная |
|
|
№ 1 |
Отверстия: №1 - М; №2 - S; №3 - S; №4 - S. Границы тела - естественные |
|
|
№2 |
Отверстия: №1 - М; №2 - S; №3 - L; №4 - S. Границы тела - измененные (с креплением пластикового объекта) |
Отверстия: №1 - S; №2 - S; №3 - М; №4 - S. Границы тела - естественные |
|
№3 |
Отверстия: №1 - М; №2 - L; №3 - М; №4 - S. Границы тела - измененные (с креплением пластикового объекта) |
Отверстия: №1 - S; №2 - М; №3 - S; №4 - S. Границы тела - естественные |
|
№ 4 |
Отверстия: №1 - М; №2 - L; №3 - М; №4 - S. Границы тела - измененные (с креплением металлического груза) |
- |
|
№ 5 |
Отверстия: №1 - М; диаметр остальных отверстий избирался случайным образом при условии, чтобы хотя бы одно было L. Границы тела - измененные (с креплением пластикового объекта или металлического груза) |
- |
Серия № 1. Ставилась задача сформировать у крыс обеих групп навык доставания приманки через отверстие № 1 (другие отверстия имели слишком маленький диаметр). О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества неуспешных попыток достижения приманки через отверстия № 2, 3 и 4.
Серия № 2. Границы тела крыс экспериментальной выборки увеличивались с помощью крепления на теменную часть головы животного цилиндрического пластикового объекта высотой 12 мм, диаметром 22 мм и весом 3 г (далее - большой груз). Объект крепился непосредственно перед началом каждой пробы и снимался после ее окончания. После увеличения границ тела животные не могли проникать головой в отверстия типа М, т. е. доставать приманку для них было возможно только через отверстия типа L. Ставилась задача выяснить, смогут ли крысы, при условии увеличения границ их тела до такого размера, что решение прежней задачи ранее выученным способом (получение приманки через отверстие № 1) окажется невозможным, модифицировать собственное поведение для достижения желаемого результата, т. е. найти новый путь через отверстие № 3. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение неуспешных попыток достижения приманки через отверстия № 1, 2 и 4.
В контрольной выборке границы тела крыс оставались неизменными, однако отверстие № 1, через которое в предыдущей серии у них был сформирован навык достижения приманки, делалось непроницаемым (S), а проницаемым (М) становилось отверстие № 3. Задача серии заключалась в том, чтобы определить, смогут ли крысы, при условии неизменности физических параметров тела, но изменении условий внешней среды найти новый путь к приманке. Полученные данные (по количеству проникновений в отверстия и времени решения экспериментальной задачи) сопоставлялись с данными, полученными на испытуемых экспериментальной выборки в этой же серии, для определения различий в специфике влияния на поведение крыс ситуации невозможности реализации ранее сформированного навыка — в первом случае (экспериментальная группа) такая ситуация возникает вследствие изменения параметров тела животного, а во втором случае (контрольная группа) такая ситуация возникает вследствие изменения параметров внешней среды.
Серия №3. Проводилась при условии, что к концу серии № 2 у испытуемых обеих выборок удалось сформировать новый навык достижениями приманки через отверстие № 3.
В экспериментальной группе ставилась задача установить, как быстро и после какого количества ошибок (неуспешных попыток достижения приманки) крысы смогут сформировать новый навык достижения приманки через отверстие № 2 при условии, что границы тела останутся увеличенными, но размер отверстия № 3 уменьшится до М, т. е. станет непроницаемым для их тела. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества неуспешных попыток достижения приманки через отверстия № 1, 3 и 4.
В контрольной выборке границы тела оставались неизменными, однако отверстие № 3, через которое в предыдущей серии у них был сформирован навык достижения приманки, делалось непроницаемым (S), а проницаемым (М) становилось отверстие № 2. Ставилась задача определить, смогут ли крысы при условии неизменности физических параметров тела, но изменении условий внешней среды найти новый путь к приманке.
Данные, полученные в экспериментальной и контрольной выборках, сопоставлялись друг с другом.
Серия № 4. Проводилась при условии, что к концу серии № 3 у испытуемых контрольной выборки удалось сформировать новый навык достижения приманки через отверстие № 2.
Серия проводилась только в экспериментальной группе. Расположение отверстий - такое же, как и в серии № 2. Границы тела крыс уменьшались до естественных. Ставилась задача: выяснить, сформируют ли крысы экспериментальной группы навык доставания приманки через отверстия типа М в ситуации, когда это стало вновь возможным благодаря уменьшению границ их головы.
Серия № 5. Проводилась только в экспериментальной группе с целью исключения возможности выбора крысой различной стратегии поведения с ориентаций на вес объекта, закрепленного у нее на голове. Границы тела для каждой пробы устанавливались случайным образом: либо они увеличивались с помощью крепления большого груза, либо на голову крысы крепился металлический груз высотой 3 мм, диаметром 12 мм и весом 3 г — данный груз не препятствовал проникновению в отверстия типа М, но весил столько же, сколько и пластиковый объект (далее - малый груз). Расположение отверстий также варьировалось случайным образом в каждой пробе (между отверстиями типов М и L), за исключением отверстия № 1, которое всегда оставалось М, а также при условии, что хотя бы одно отверстие являлось L. Ставилась задача выяснить, как часто крысы будут совершать попытки проникновения в отверстия типа М при креплении малого и большого грузов и будут ли выявлены различия, свидетельствующие об ориентации животных на вес груза.
Процедура модификации границ тела крыс. В теменную кость черепа крыс (экспериментальной группы) монтировался винт из нержавеющей стали шляпкой вниз. В результате данной операции после зашивания шва часть винта с резьбой выступала над скальпом на высоту 3 мм. К винту в дальнейшем крепились большой и малый грузы (рис. 2). Данная операция проводилась в соответствии с современными принципами биоэтики (Beauchamp, Childress, 2001) и являлась необходимой, так как неинвазивные техники не позволяли достаточно устойчиво закрепить инородный объект на теле крысы. Необходимо отметить, что при креплении винта в черепе не делалось сквозного отверстия, что гарантировало целостность мозговых структур животного.

Рис 2. Крыса из экспериментальной группы с закрепленным на голове большим грузом
Операция проводилась за 14 дней до начала экспериментальной серии № 1. Объективным критерием того, что проведенная операция не оказала влияния на специфику соотнесения крысами собственного тела с объектами внешней среды и, соответственно, на их схему тела, должно было являться отсутствие статистических различий в решении задачи между испытуемыми контрольной и экспериментальной выборок по критерию времени и количества проникновений в различные типы отверстий.
Аппаратура. Поведение животных в процессе эксперимента фиксировалось с помощью видеокамеры (Sony HDR-CX405), для фиксации временных интервалов использовался таймер видеозаписи данной камеры.
Регистрируемые показатели:
• время (в секундах) решения экспериментальной задачи в каждой пробе в сериях № 1-3 в обеих группах регистрировалось от момента помещения крысы в пусковую камеру до момента достижения ею приманки через одно из отверстий;
• количество попыток проникновения (успешных и неуспешных) в различные типы отверстий отдельно для каждого отверстия в каждой пробе и отдельно для каждой крысы в каждой пробе во всех сериях в обеих группах. За одну попытку проникновения считались ситуации, когда крыса погружала голову в отверстие хотя бы на несколько миллиметров.
Особое внимание было уделено неуспешным попыткам проникновения в отверстия типа М, совершенным крысами экспериментальной группы с увеличенными границами тела в сериях № 2, 3 и 5, что объяснялось их существенным разнообразием. В ходе пилотажных исследований было установлено, что эти попытки имеют значительный разброс по продолжительности, а также имеют разнообразные поведенческие проявления.
В одних случаях крыса максимально глубоко проникала головой в отверстие, упираясь в него большим грузом и совершая буксующие движения лапами в сторону отверстия. Длительность таких попыток составляла 5-8 с. Данное поведение, на наш взгляд, свидетельствовало о стремлении крысы достать приманку, игнорируя тот факт, что границы головы не позволяют ей это сделать. Эти попытки мы считали длительными попытками.
В других случаях крыса только проникала головой в отверстие (при этом проникновение зачастую оказывалось не максимально глубоким) и затем вынимала голову назад; буксующие движения лапами отсутствовали. Длительность таких попыток составляла 0,5-1,5 с. Как мы полагаем, в данном случае крыса учитывала тот факт, что границы ее головы не позволяют ей достать приманку через это отверстие. Само проникновение осуществлялось крысой лишь для того, чтобы определить размер отверстия. Эти попытки были названы нами краткими попытками.
Соответственно, для дальнейшего разделения попыток проникновения на длительные и краткие фиксировались следующие показатели неуспешных попыток проникновения в отверстия типа М, совершенных испытуемыми экспериментальной выборки в каждой пробе серий № 2, 3 и 5:
• длительность каждой попытки проникновения (в секундах), считавшаяся от момента, когда крыса погружала голову в отверстие хотя бы на несколько миллиметров, до момента, когда крыса полностью вынимала голову из отверстия;
• наличие или отсутствие буксующих движений лапами, совершаемых крысой при попытке проникновения в отверстие.
На основе этих критериев все вышеописанные попытки были дифференцированы на длительные и краткие и отдельно зарегистрированы в каждой пробе серий № 2, 3 и 5 у испытуемых экспериментальной группы с увеличенными границами головы (крепление большого груза).
Гипотеза исследования могла считаться подтвержденной при получении следующих фактов:
• уменьшение количества длительных попыток проникновения в сопоставлении с краткими попытками проникновения у испытуемых экспериментальной группы при переходе от серии № 2 к серии № 3;
• в серии № 3 отсутствие достоверных различий в распределении по количеству попыток проникновения в различные номера отверстий между испытуемыми контрольной и экспериментальной групп;
• в серии № 3 отсутствие достоверных различий между испытуемыми контрольной и экспериментальной групп во временных интервалах, затраченных на решение экспериментальной задачи.
Такие факты дали бы основание утверждать, что крысы экспериментальной выборки не просто научились не проникать в отверстия типа М, располагающиеся в определенных частях экспериментальной установки (в серии № 2), но и в целом научились воспринимать отверстия данного типа как «непроницаемые» для своих новых размеров тела, даже в случае расположения этих отверстий в других частях экспериментальной установки (в серии № 3). Соответственно, данный факт свидетельствовал бы о том, что крысы модифицировали схему собственного тела. Также в случае модификации схемы тела к концу серии № 2 у испытуемых экспериментальной выборки в серии № 3 должны будут отсутствовать значимые различия между контрольной и экспериментальной группами в распределении попыток проникновения в различные номера отверстий, а также во временной динамике формирования нового навыка. Результаты серий № 4 и 5 были необходимы для установления того, насколько гибко крысы способны перестраивать схему собственного тела в зависимости как от изменений характеристик внешней среды, так и от изменений границ собственного тела.
Переменные.
Независимые переменные:
• границы тела крыс: естественные (естественными считались границы тела испытуемых контрольной группы, а также экспериментальной группы в отсутствие закрепленного на их головах большого или малого грузов), измененные с помощью большого груза, измененные с помощью малого груза;
• диаметр отверстий, соединявших центральный отсек с обрамляющими отсеками.
Зависимые переменные:
• время решения экспериментальной задачи в каждой пробе;
• количество успешных и неуспешных попыток проникновения в различные типы отверстий (в том числе, количество длительных и кратких попыток у испытуемых экспериментальной группы в сериях № 2,3 и 5).
Математический анализ осуществлялся с помощью программы Statistica 8.
Результаты
Серия № 1. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 3) (в экспериментальной группе: Тест Вил- коксона Т=0; Z=2.52; п=8; р<0.05, в контрольной группе: Т=0; Z=2.52; п=8; р<0.05). Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 1834 с (SD=180.2), в контрольной группе - 1795 с (SD=154.9). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 7 с (SD=1.1), в контрольной группе - 8 с (SD=1.4). По показателям времени решения экспериментальной задачи испытуемые экспериментальной и контрольной групп не отличались друг от друга (Тест Манна-Уитни U=26.5; n4=8; n2=8; р>0.05).
В экспериментальной и контрольной группах к концу серии крысы стали чаще совершать попытки проникновения в отверстия № 1 и реже - в отверстия № 2, 3 и 4 (рис. 3). В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 1 составляла 21%, на последних пяти пробах - 100%; х2=89.22; df=3; р<0.01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 1 составляла 22%, на последних пяти пробах - 100%; /2=86.60; df=3; р<0,01. По общим соотношениям количества попыток проникновения в отверстия различных номеров достоверных отличий у испытуемых двух групп не обнаружено (х2=0.04; df=3; р>0.05).
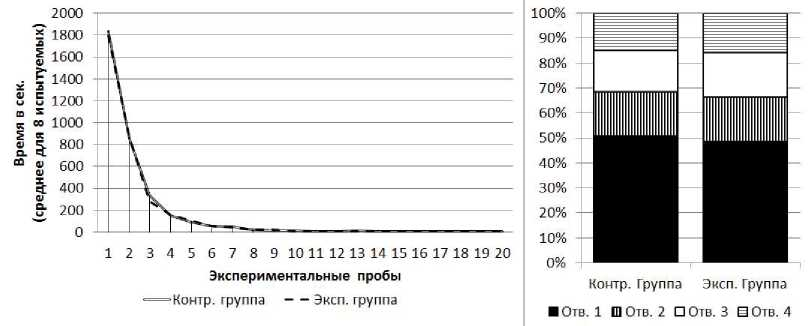
Рис 3. Слева - кривые научения контрольной и экспериментальной групп в серии № 1. Справа-распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 1
Серия №2.У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (в экспериментальной группе: Т=0; Z=2.52; п=8; р<0.01, в контрольной группе: Т=0; Z=2.52; п=8; р<0.01). Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 524 с. (SD=57.4), в контрольной группе - 52 с. (SD=10.3). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 17 с (SD=2.5), в контрольной группе - 17 с. (SD=2.9). На решение экспериментальной задачи крысы обеих групп в серии № 2 затратили меньше времени, нежели в серии № 1 (в экспериментальной группе: Тест Вилкок- сона Т=0; Z=2.52; п=8; р<0.05, в контрольной группе: Т=0; Z=2.52; п=8; р<0.05). При этом на решение экспериментальной задачи испытуемые экспериментальной группы затратили больше времени, нежели испытуемые контрольной группы (U=0; nt=8; n2=8; р<0.01).
В экспериментальной и контрольной группах к концу серии крысы стали чаще совершать попытки проникновения в отверстия № 3 и реже - в отверстия № 1, 2 и 4. В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 3 составляла 22%, на последних пяти пробах - 50%; /2=25.19; df=3; р<0.01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 3 составляла 40%, на последних пяти пробах - 50%; х2=16.02; df=3; р<0.01 (рис. 4). Доля неуспешных попыток проникновения в отверстие № 1 (через это отверстие испытуемые достигали приманку в прошлой серии) за все 20 проб серии № 2 у испытуемых экспериментальной группы составляет 58%, а у испытуемых контрольной группы - 47%. Крысы экспериментальной группы чаще совершали попытки проникновения в отверстие № 1, нежели испытуемые контрольной группы (х2=8.65; df=3; р<0.05).
Отдельно следует отметить, что даже к концу серии № 2 при сокращении времени решения задачи, а также уменьшении общего количества неуспешных попыток достижения приманки все испытуемые (обеих выборок) в начале каждой пробы совершали попытку проникновения в отверстие № 1 и только после этого направлялись к отверстию № 3, чтобы достать приманку.
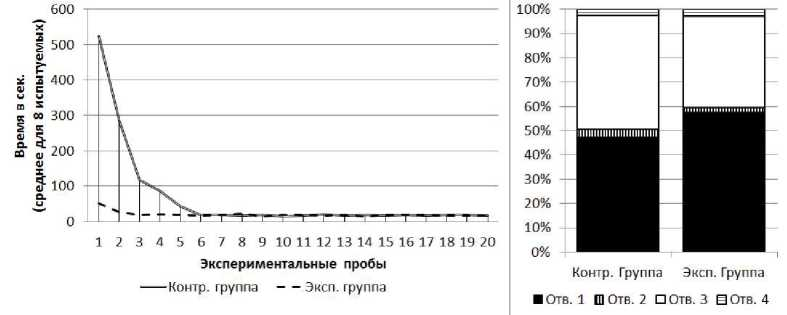
Рис 4. Слева - кривые научения контрольной и экспериментальной групп в серии № 2. Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 2
Серия № 3. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 5) (в экспериментальной группе: Т=0; Z=2.52; п=8; р<0.01, в контрольной группе: Т=0; Z=2.52; п=8; р<0.01). Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 48 с (SD=12.2), в контрольной группе - 51 с (SD=9.8). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 17 с (SD=2.2), в контрольной группе - 18 с (SD=3.1). На решение экспериментальной задачи в серии № 3 крысы контрольной группы затратили столько же времени, сколько и в серии № 2 (Тест Вилкоксона Т=4; Z=1.34; п=8; р>0.05), а крысы экспериментальной группы в серии № 3 затратили меньше времени, нежели в серии № 2 (Тест Вилкоксона Т=0; Z=2.52; п=8; р<0.05). При этом, в серии № 3 на решение экспериментальной задачи крысы обеих групп затратили одинаковое количество времени (U=27; nt=8; n2=8; р>0.05).
В экспериментальной и контрольной группах к концу серии крысы стали чаще совершать попытки проникновения в отверстия № 2 и реже - в отверстия № 1, 3 и 4. В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 2 составляла 38%, на последних пяти пробах - 50%; х2=22.02; df=3; р<0.01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 3 составляла 40%, на последних 5 пробах - 50%; х2=18.00; df=3; р<0.01 (рис. 5). Общие соотношения количества попыток проникновения в отверстия различных номеров у испытуемых двух групп не имеют достоверных отличий друг от друга (%2=2.78; df=3; р>0.05).
В серии № 3, как и в предыдущей серии, в начале каждой пробы все испытуемые обеих групп совершали одну неуспешную попытку проникновения в отверстие № 1.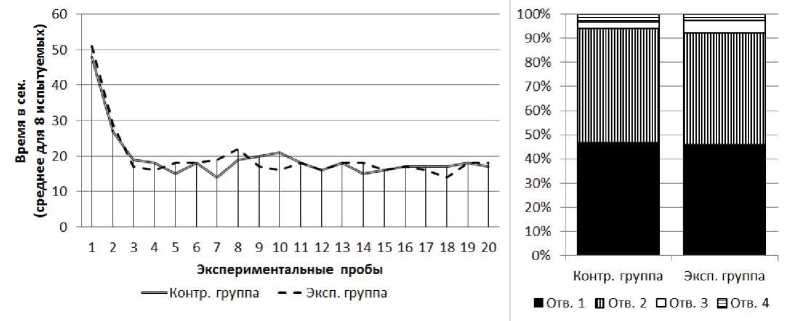
Рис 5. Слева - кривые научения контрольной и экспериментальной групп в серии № 3. Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 2
Серия № 4. После уменьшения тела до естественных размеров крысы экспериментальной группы осуществляли достижение приманки через отверстие типа L только на первых четырех пробах данной серии. Общее количество достижений приманки на первых пяти пробах составило: через отверстие L - 16, через отверстие М — 24; на последних пяти пробах: через отверстие L — 0, через отверстие М - 40 (различия достоверны %2=20.00; df= 1; р<0.01). При этом все достижения приманки через отверстия типа М осуществлялись через отверстие № 1, хотя отверстие № 3 имело такой же диаметр. Также отметим, что, как и в предыдущих двух сериях, здесь в начале каждой пробы крысы осуществляли неуспешную попытку проникновения в отверстие №1.
Серия № 5. При креплении малого груза крысы экспериментальной группы достигали приманку через отверстия типа М в 85 случаях, через отверстия типа L - в 3 случаях. При креплении большого груза крысы экспериментальной группы достигали приманку через отверстия типа М в 0 случаях, через отверстия типа L - в 72 случаях. Различия достоверны (х2=148.36; df=l; р<0.01). Также отметим, что, как и в предыдущих трех сериях, здесь в начале каждой пробы крысы осуществляли неуспешную попытку проникновения в отверстие № 1.
Количество длительных и кратких неуспешных попыток проникновения. Длительные и краткие попытки проникновения в отверстия типа М подсчитывались у испытуемых экспериментальной группы в тех пробах, когда границы их тела увеличивались с помощью большого груза (в сериях № 2,3 и 5). Всего в данных условиях было зафиксировано 496 попыток проникновения в отверстия типа М. Результаты кластерного анализа представлены в табл. 2 и 3.
Таблица 2
Группа •«длительных попыток проникновения*, выделенная в результате кластерного анализа методом k-средних (в таблице приведены данные после проведения стандартизации)
|
Переменные |
Mean |
SD |
DX |
|
Длительность попытки проникновения |
2.81 |
0.55 |
0.30 |
|
Наличие/отсутствие буксующих движений лапами |
-2.92 |
<0.01 |
<0.01 |
Таблица 3
Группа -«кратких попыток проникновения*, выделенная в результате кластерного анализа методом k-средних (в таблице приведены данные после проведения стандартизации)
|
Переменные |
Mean |
SD |
DX |
|
Длительность попытки проникновения |
-0.33 |
0.22 |
0.05 |
|
Наличие/отсутствие буксующих движений лапами |
0.34 |
<0.01 |
<0.01 |
К группе длительных попыток проникновения было отнесено 52 попытки, средняя длительность попытки проникновения - 6,75 с (SD=1.01), в ходе всех попыток проникновения крысы совершали буксующие движения лапами. К группе кратких проникновений было отнесено 444 попытки, средняя длительность попытки проникновения - 0,96 с (SD=0.41), в ходе данных попыток не было зафиксировано ни одного случая совершения крысой характерных буксующих движений лапами.
Все 52 длительные попытки проникновения были зафиксированы в течение первых пяти проб серии № 2. Краткие попытки проникновения наблюдались в ходе каждой пробы в течение всех трех вышеуказанных серий. В том числе все попытки проникновения в отверстие № 1, совершаемые крысами экспериментальной группы в начале каждой пробы, начиная с серии № 2, являлись краткими. Соответственно, в серии № 2 испытуемые совершили достоверно больше длительных попыток проникновения, нежели в серии № 3 ( 2=42.89; df=l; р<0.01) и серии № 5 (х2=18.19; df= 1; р<0.01).
Обсуждение результатов
Отсутствие в серии № 1 различия между контрольной и экспериментальной группами по времени решения задачи и количеству проникновений в различные типы отверстий свидетельствует о том, что подготовительная операция (для увеличения границ тела испытуемых в следующих сериях), проведенная на крысах экспериментальной группы, не оказала существенного влияния на специфику соотнесения ими собственного тела с объектами внешней среды. Иными словами, данная операция не привела к модификации схемы тела крыс экспериментальной группы.
Сокращение временных интервалов решения задачи и количества неуспешных попыток достижения приманки к концу серий № 1-3 у испытуемых обеих групп свидетельствует о формировании у крыс нового навыка достижения приманки в каждой из первых трех серий.
В серии № 2 животные экспериментальной группы тратили больше времени на решение задачи и совершали больше неуспешных попыток достижения приманки через отверстие № 1, нежели испытуемые контрольной группы. Это свидетельствует о том, что для крысы формирование нового навыка, связанного с изменением характеристик собственного тела, является более трудоемкой процедурой, нежели формирование нового навыка, обусловленного изменениями характеристик внешних объектов.
Мы заключаем, что главная гипотеза исследования была подтверждена на основании следующих данных.
Во-первых, испытуемые экспериментальной группы совершали длительные неуспешные попытки проникновения в отверстия типа М только в течение первых пяти проб серии № 2 (т. е. достоверно чаще, нежели в сериях № 3 и № 5). Как уже отмечалось, мы полагаем, что эти попытки свидетельствуют об игнорировании крысой увеличенных границ своего тела, т. е. о том, что крыса взаимодействует с внешним миром, используя старую схему собственного тела, нерелевантную тем модификациям, которые было произведены с ее головой. Уменьшение длительных попыток свидетельствует о том, что крыса научается учитывать изменения границ своего тела, т. е. модифицирует схему собственного тела. Более того, поскольку формирование навыка происходило довольно быстро — за пять проб (каждая крыса совершила в среднем 6,5 длительных попыток проникновения) - и более не наблюдалось в сериях № 3 и № 5, мы полагаем возможным расценить приобретение данного опыта как инсайт (Кёлер, 1930). В противовес этому, краткие попытки проникновения, как мы полагаем, осуществляются не с целью достичь приманку, но с ориентировочной целью - оценить размеры отверстия и, возможно, границы собственного тела. В процессе краткой попытки проникновения крыса соотносит габариты собственной головы с диаметром отверстия, очевидно, опираясь, главным образом, на кинестетические ощущения, так как зрение у этих животных развито слабо. Соответственно, краткие попытки (и их количество) не свидетельствуют о том, что крыса игнорирует границы собственного тела, используя старую схему тела.
Во-вторых, в серии № 3 отсутствуют достоверные различия между испытуемыми контрольной и экспериментальной групп по времени решения задачи и количеству проникновений в отверстия различных типов. Этот факт мы объясняем тем, что в серии № 3 крысы экспериментальной группы формируют новый навык, основываясь на уже модифицированной схеме собственного тела. Соответственно, данный навык складывается быстрее, чем у них же в предыдущей серии (когда нужно было изменять старую схему своего тела) - с такой же скоростью (по критерию времени и количества неуспешных попыток достижения приманки), что и у крыс контрольной группы в этой же серии № 3.
Таким образом, мы констатируем, что при изменении границ тела крыс в сторону ограничения/препятствования осуществлению животными ранее выученного поведения, эти животные способны модифицировать схему своего тела, что выражается в том, что они перестраивают свое прежнее поведение с учетом изменений, которые претерпело их тело.
Результаты, полученные в серии № 4, свидетельствуют, что при уменьшении границ тела крысы способны переучиваться, снова модифицируя схему собственного тела и достигая приманку кратчайшим путем через отверстие № 1 - типа М. Более того, факты, полученные в серии № 5, позволяют заключить, что крысы могут гибко менять свое поведение в зависимости от того, закреплен на их голове большой груз, препятствующий достижению приманки через отверстия типа М, или малый груз, не препятствующий осуществлению этой операции. Очевидно, это также свидетельствует о способности крыс гибко перестраивать схему собственного тела, при этом данная перестройка базируется не на параметрах веса груза, так как по этому параметру большой и малый груз были идентичны.
Отдельного обсуждения заслуживает тот факт, что испытуемые обеих групп, начиная с серии № 2, в начале каждой пробы совершают попытку проникновения в отверстие № 1 даже в случае, если у них уже сформирован навык. Мы полагаем, что в результате подобной операции проникновения крысы осуществляют оценку размеров отверстия, а также границ собственного тела (в случае с крысами экспериментальной группы). То обстоятельство, что крысы осуществляли эту операцию именно с помощью отверстия № 1, вероятно, связано, во-первых, с его близостью (так как в начале каждой пробы запускали именно в отсек с отверстием № 1), во-вторых, с ригидностью поведения: сохранностью самого раннего навыка достижения приманки через данное отверстие в серии № 1.
В ранних исследованиях, проводившихся на членистоногих (в частности, тараканах Periplaneta americana (Хватов, 2011)), в ходе которых у животных экспериментальной группы также увеличивали границы тела, а для контрольной группы изменялась схема отверстий в экспериментальной установке, были получены существенно иные результаты. Во- первых, неуспешные попытки проникновения в ранее проницаемые отверстия у этих животных не удалось разделить на длительные и краткие. Во-вторых, членистоногие, как при изменении границ тела, так и при изменении размера отверстий, демонстрировали одинаковое поведение (по временным параметрам формирования навыка и по соотношению количества попыток проникновения в различные типы отверстий). В-третьих, членистоногие не переносили ранее приобретенный опыт, связанный с изменениями границ их тела, из одной серии в другую - в каждой следующей серии навык формировался у них «с нуля»: животные экспериментальной группы совершали такое же количество неуспешных попыток проникновения в отверстия типа М, как и в предыдущей серии.
В аналогичных экспериментах, проводившихся на змеях (Хватов и др., 2015b), было установлено, что эти животные, аналогично членистоногим, не переносят приобретенный опыт, связанный с изменениями границ их тела, в новую ситуацию. Однако, в отличие от членистоногих, в поведении змей выделялись краткие и длительные попытки проникновения в отверстия типа М. В рамках одной пробы длительные попытки проникновения обнаруживались чаще в начале, а краткие - в конце пробы, что, по мнению авторов, свидетельствовало о возможности модификации схемы тела, хотя и в весьма узких временных рамках (в пределах отдельной пробы).
В аналогичных экспериментах, проводившихся на ящерицах (Бурыченкова, Хватов, 2015), было установлено, что эти животные все же способны переносить приобретенный опыт, связанный с изменениями границ их тела, в новую ситуацию. Однако это выражается в виде постепенного уменьшения количества попыток проникновения в непроницаемые отверстия типа М в последующих сериях (при изменении расположений отверстий в экспериментальной установке), но не в их полном отсутствии, как это было обнаружено у крыс в настоящем эксперименте. Данный факт можно объяснить тем, что у ящериц модификация схемы тела происходила не путем инсайта (как у крыс), а путем постепенного научения.
С позиции разрабатываемой нами концепции самоотражения животных и человека (Хватов, 2010; 2014), полученные в результате сравнения данные можно проинтерпретировать следующим образом: в психике членистоногих схема тела в качестве самостоятельной автономной структурной единицы еще не выделяется. Однако осуществлять какую бы то ни было поведенческую активность без учета характеристик собственного тела невозможно. У членистоногих отражение характеристик собственного тела существует в неразрывном единстве с отражением внешних условий (схемы расположения отверстий в экспериментальной установке). Можно утверждать, что субъективно они воспринимают лишь собственную «неспособность» проникнуть в отверстие, но не воспринимают, чем обусловлена эта неспособность: изменениями их собственного тела или изменениями внешней среды. Поэтому, сталкиваясь с такой «неспособностью», членистоногие полностью перестраивают психическое отражение собственного co-бытия с внешним миром вместе с восприятием собственного тела и восприятием внешней среды, т. е. выстраивают полностью новый навык - «с нуля».
У пресмыкающихся - во всяком случае, у некоторых представителей - обнаруживается большая автономия (и, соответственно, возможность для более гибкой трансформации) схемы собственного тела от схемы внешней среды в рамках целостного психического отражения организмом собственного co-бытия с внешней средой. Это проявляется в том, что животные уже способны модифицировать схему собственного тела путем научения, постепенно обобщая опыт собственного взаимодействия с объектами внешнего мира в различных ситуациях.
У крыс схема тела уже обособляется в самостоятельный когнитивный конструкт и, будучи сформирована в рамках одного события (в рамках связи с определенными характеристиками внешней среды), может быть экстраполирована за его пределы и включена в рамки других событий (в связи с другими характеристиками внешней среды). Соответственно, у крыс, в сравнении с членистоногими и пресмыкающимися, психический образ имеет большую дифференцированность и гибкость организации, что позволяет использовать элементы ранее сформированных навыков для выстраивания новых и благодаря этому экономить время и совершать меньше ошибок. Это соответствует общему представлению о направлениях эволюции психики у представителей разных эволюционных таксонов (Хватов, 2012). Однако в отличие от животных, осуществляющих ориентацию во внешней среде преимущественно на основе визуальной информации (в частности - приматов), крысы соотносят характеристики собственного тела с характеристиками объектов внешней среды преимущественно на основе кинестетической информации. Мы полагаем, что, визуальная информация в этом случае носит лишь вспомогательный характер. С учетом данных, полученных на других млекопитающих (как авторским коллективом, так и другими исследователями - подробнее см.: Хватов, 2014), можно предположить, что многие млекопитающие обладают сложно организованной схемой собственного тела; возможно, у них даже складываются определенные представления о себе, однако роль визуальной информации в построении этих представлений относительно невелика. Данный тезис указывает на неприменимость некоторых классических методик (например, «Теста с зеркалом») на нечеловекообразных видах животных (англ, «non-human species»).
Выводы
На основе проведенного исследования мы констатируем, что при изменении границ тела крыс в сторону ограничения/препятствования осуществлению ими ранее выученного поведения, животные модифицируют схему своего тела, что выражается в перестройке животными своего прежнего поведения с учетом изменений собственного тела. Более того, уже обученные крысы способны перестраивать схему своего тела в зависимости от того, увеличены ли физические границы их тела или нет.
* Хватов И. А. Кандидат психологических наук, доцент, кафедра общей психологии и истории психологии, Московский гуманитарный университет; Московский институт психоанализа. E-mail: ittkrot 1 ©gmail.com
* * Соколов А.Ю. Кандидат биологических наук, старший научный сотрудник, АНО Лаборатория- студия -«Живая Земля». E-mail: apophis-king@mail.ru
* **Харитонов А.Н. Кандидат психологических наук, научный сотрудник, Институт психологии РАН; ведущий научный сотрудник, Центр экспериментальной психологии МГППУ. E-mail: ankhome47@list.ru ****КуличенковаК.Н. Аспирант, биологический факультет, Московский государственный университет имени М.В. Ломоносова. E-mail: koulitchenkova@gmail.com
Литература
- Барабанщиков В. А. Восприятие и событие. СПб.: Алетейя, 2002.
- Бурыненкова Д. С., Хватов И. А. Особенности восприятия границ собственного тела сцинками Tili- qua gigas // Психология - наука будущего: Материалы VI Международной конференции молодых ученых / Под ред. А.Л. Журавлева, Е.А. Сергиенко. М.: Институт психологии РАН, 2015. С. 78-82.
- Кёлер В. Исследование интеллекта человекообразных обезьян. М.: Изд-во Комакадемии, 1930.144 с.
- Хватов И. А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011. Т. 4. № 1. С. 28-39.
- Хватов И.А. Главные направления эволюции психики в контексте онтологического и диффе- ренционно-интеграционного подходов. Часть 1 [Электронный ресурс] // Психологические исследования. 2012а. № 1 (21). С. 1. URL: http://psystudy.ru/index.php/num/2012nl-21/622-khvatov21.html (дата обращения: 22.09.2014). doi: 0421200116/0001
- Хватов И. А. Эволюция самоотражения животных и человека в контексте дифференционно- интеграционного подхода // Дифференционно-интеграционная теория развития. Кн. 2. / Сост. и ред. Н.И. Чуприкова, Е.В. Волкова. М.: Языки славянских культур, 2014. С. 343-360.
- Хватов И. А., Харитонов А.Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. Т. 5. № 3. С. 96-107.
- Хватов И. А., Харитонов А. Н., Соколов А. Ю. Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Giyllus assimilis // Экспериментальная психология. 2013. Т. 6. № 4. С. 79-95.
- Хватов И.А., Соколов А.Ю., Харитонов А.Н. Схема собственного тела у змей Lampropeltis triangulum campbelli // Экспериментальная психология. 2015. Т. 8. №2. С.119-138. doi:10.17759/ exppsy.2015080209
- Хватов И.А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Восприятие собственного тела у крыс // Естественно-научный подход в современной психологии / Отв. ред. В. А. Барабанщиков. М.: Институт психологии РАН, 2014а. С. 326-332.
- Хватов И. А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Методика изучения схемы тела у мелких млекопитающих // Экспериментальная психология. 2014b. Т. 7. № 3. С. 137-144.
- Beauchamp T.L. Childress J.F. Principles of Biomedical Ethics. N.Y.: Oxford University Press, 5th ed., 2001.454 p.
- Barnes C.A. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat //Journ. Comp. Physiol. Psychol. 1979. Vol. 93. № 1. P. 74-104.
- Berti A. Frassinetti F. When Far Becomes Near: Remapping of Space by Tool Use //Journ. of Cognitive Neuroscience. 2000. Vol. 12. № 3. P. 415-420. doi:10.1162/089892900562237
- Carlson T. Alvarez G. Wu Daw-an Verstraten F.A.J. Rapid Assimilation of External Objects Into the Body Schema// Psychological Science. 2010. Vol. 21. № 7. P. 1000-1005. doi:10.1177/0956797610371962
- Costantini M. Frassinetti F. Maini M. Ambrosini E. Gallese V. Sinigaglia C. When a laser pen becomes a stick: remapping of space by tool-use observation in hemispatial neglect // Experimental Brain Research. 2014. Vol. 232. № 10. P. 3233-3241.
- Gallagher S. Cole J. Body Schema and Body Image in a Deafferented Subject //Journ. of Mind and Behavior. 1995. Vol. 16. P. 369-390. doi: 10.1068/p7027
- Garbarinia F., Fossataroa C., Bertia A., Gindria P., Romanod D., Piaa L., GattafF., Maravitad A., Neppi-Modona M. When your arm becomes mine: Pathological embodiment of alien limbs using tools modulates own body representation //Neuropsychologia. 2015. Vol. 70. P. 402-413. doi:10.1016/j.neuropsychologia.2014.11.008
- Giglia G., Pia L., Folegatti A., Puma P., Fierro B., Cosentino G., Berti A., Brighina F. Far Space Remapping by Tool Use: A rTMS Study Over the Right Posterior Parietal Cortex // Brain Stimulation. 2015. Vol. 8. № 4. P. 795-800. doi: https://doi.org/10.1016/j.brs.2015.01.412
- Gozli D. G., Brown L.E. Agency and Control for the Integration of a Virtual Tool into the Peripersonal Space//Perception. 2011. Vol. 40. № 11. P. 1309-1319.
- Johnson-Frey S. The neural bases of complex tool use in humans // Trends in Cognitive Sciences. 2004. Vol. 8. № 2. P. 71-78. doi:10.1016/j.tics.2003.12.002
- Maravita A., Iriki A. Tools for the body (schema) // Trends in Cognitive Sciences. 2004. Vol. 8. № 2. P. 79-86.
- Moeller B. Zoppke H. Frings C. What a car does to your perception: Distance evaluations differ from within and outside of a car // Psychonomic Bulletin & Review. 2015. doi: 10.3758/cl3423-015-0954-9
- Morris R. Developments of a water-maze procedure for studying spatial learning in the rat //Journal of neuroscience methods. 1984. Vol. 11. № 1. P. 47-60.
- Ritchie J.B. Carlson T.A. Tool Integration and Dynamic Touch // Psychological Science. 2013. doi: 10.1177/0956797612459768
Информация об авторах
Метрики
Просмотров
Всего: 5067
В прошлом месяце: 9
В текущем месяце: 5
Скачиваний
Всего: 2223
В прошлом месяце: 5
В текущем месяце: 0