Экспериментальная психология
2018. Том 11. № 4. С. 5–16
doi:10.17759/exppsy.2018110401
ISSN: 2072-7593 / 2311-7036 (online)
Особенности научения жаб Bufo viridis в поведении, требующем учета границ собственного тела
Аннотация
Общая информация
Ключевые слова: научение, схема тела, земноводные, жабы, самоотражение, воплощенное познание
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2018110401
Финансирование. Исследование поддержано грантом Совета по грантам при Президенте Российской федерации № МК-2766.2017.6.
Для цитаты: Хватов И.А., Желанкин Р.В. Особенности научения жаб Bufo viridis в поведении, требующем учета границ собственного тела // Экспериментальная психология. 2018. Том 11. № 4. С. 5–16. DOI: 10.17759/exppsy.2018110401
Полный текст
Традиционно хладнокровные позвоночные считаются ограниченными с точки зрения своих когнитивных способностей в сравнении с млекопитающими и птицами. Однако современные данные показывают, что большая часть поведенческого репертуара рептилий и амфибий является не врожденной, а приобретается в форме либо облигатного, либо факультативного научения. При взаимодействии с окружающей средой эти животные демонстрируют сложные коммуникативные навыки, способность ориентироваться в сложных лабиринтах, заботу о потомстве и даже игру (Burghardt, 2013). В частности, некоторые авторы полагают, что именно у земноводных в ходе эволюции складываются поведенческие модели пространственной ориентации, характерные для амниот, в частности, теплокровных позвоночных (Daneri et al., 2011). Таким образом, изучение психических процессов земноводных является перспективной исследовательской задачей для сравнительной психологии, решение которой необходимо для более глубокого понимания особенностей психической организации у позвоночных и животных в целом.
Существуют данные о способности жаб обучаться в T-образном лабиринте (Brattstrom, 1990). При этом в большинстве работ указывается, что ключевым ориентиром при формировании навыка являются обонятельные сигналы (Ефанова и др. 2007). Также существуют данные, свидетельствующие о способности этих земноводных обучаться с опорой на визуальную стимуляцию (Daneri et al., 2011). Однако ориентиры в виде меток играют второстепенную роль в сравнении с пространственными ориентирами окружающей среды (Sotelo et al., 2015). Так, например, саламандры обучаются ориентации в Т-образном лабиринте, хотя и не способны обучаться с опорой на визуальные ориентиры (Kundey et al., 2016).
Цель настоящей работы — изучение способности зеленой жабы Bufo viridis формировать навыки пространственной ориентации с опорой на тактильные ощущения, возникающие в процессе взаимодействия тела животного и внешних объектов.
Для регуляции своего поведения в ходе взаимодействия с объектами окружающей среды любому животному необходимо воспринимать как физические характеристики этих объектов (в частности, их формы и границы), так и физические характеристики собственного тела — схему тела. Схема тела — это модель собственного тела как единого целого, включающая также совокупность представлений о физических характеристиках своего тела (его границах, весе, плотности и т. д.) и его отдельных частей, позволяющая осуществлять и планировать различные движения (Gallagher, Cole, 1995; Morasso et al., 2015). Схема представляет собой сложный ментальный конструкт и строится на основе информации различных модальностей: тактильной, аудиальной, визуальной, проприорецептивной (Gallagher, Cole, 1995). Схема тела тесно связана с восприятием размерности окружающих объектов, формируется и существует в процессе соотнесения субъектом собственных характеристик с характеристиками окружающих объектов (там же). В отличие от образа тела, схема тела является неосознаваемым феноменом (там же).
Большая часть исследований схемы тела проводилась на людях (Gallagher, Cole, 1995). В научной литературе существует лишь несколько исследований схемы тела млекопитающих (Johnson-Frey, 2004; Maravita, Iriki, 2004). Схема тела представителей других видов позвоночных не исследовалась.
Гипотезы исследования:
1. Жабы способны учитывать естественные границы собственного тела при взаимодействии с внешними объектами.
2. При экстренном изменении границ тела жаб таким образом, что эти изменения будут препятствовать осуществлению животным ранее выученного поведения, животные смогут осуществить модификацию схемы своего тела, что будет выражаться в отсутствии попыток проникновения в отверстия, слишком маленькие для их увеличенных тел.
Методика исследования
Испытуемые животные: 12 особей зеленой жабы Bufo viridis, пойманные на воле. Животные были разделены на 2 группы: контрольную (6 особей) и экспериментальную (6 особей). Выбор экспериментальных животных был обусловлен тем, что в естественной среде зеленые жабы часто прячутся в ямах и норах, которые редко роют самостоятельно (Кузьмин, 2012).
Оборудование. Экспериментальная установка представляла собой стеклянный ящик квадратной формы со стороной 700 мм и стенками высотой 500 мм, разделенный перегородкой на два отсека. В перегородке имелось три отверстия, равно отстоящие друг от друга. Диаметр отверстий мог варьироваться. Отсек № 1 являлся пусковой камерой, боковые стенки отсека были окрашены в белый цвет; над отсеком располагалась лампа (UVB200 25 ВТ), уровень освещенности составлял 300 люкс. Отсек № 2 являлся камерой влажности, на полу располагался влажный субстрат, стенки отсека (включая перегородку) были окрашены в черный цвет, сверху отсек закрывался светоизолирующей крышкой.
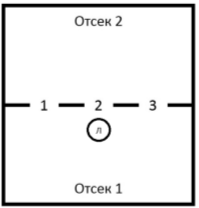
Рис. 1. Схема экспериментальной установки.
Отверстия обозначены номерами, Л — место расположения лампы
Независимые переменные в экспериментах:
• границы объектов внешней среды — диаметр отверстий в экспериментальной установке;
• границы тела животного.
В эксперименте использовалось два типа круглых отверстий в перегородке между отсеками:
1) малое отверстие диаметром 15 мм (далее — S) — в такое отверстие могла проникнуть лапа жабы или кончик ее морды, но не все животное целиком;
2) среднее отверстие диаметром 40 мм (далее — M) — достаточно крупное для проникновения через него тела жабы с естественными границами;
3) большое отверстие диаметром 70 мм (далее — L) — в такое отверстие могла проникнуть жаба с увеличенными границами тела.
Границы тела жаб увеличивались с помощью одевания на них специальных попонок, на верхней части которых крепился пластиковый цилиндрический объект высотой 15 мм и диаметром 40 мм. Попонка одевалась на испытуемых экспериментальной группы перед началом серии 2.
• время решения экспериментальной задачи в каждой пробе;
• количество успешных/неуспешных попыток проникновения в различные типы отверстий.

Рис. 2. Жаба с увеличенными границами тела
Соответственно, в ходе эксперимента фиксировались следующие показатели:
• время (в секундах) решения экспериментальной задачи в каждой пробе, отсчет которого в обеих группах производился от момента помещения жабы в центр отсека № 1 и до момента проникновения в отсек № 2 через одно из отверстий;
• количество попыток проникновения (успешных и неуспешных) в различные типы отверстий отдельно для каждого отверстия в каждой пробе и отдельно для каждой жабы в каждой пробе во всех сериях, в обеих группах. Неуспешными попытками считались такие, при которых жаба погружала голову в одно из отверстий, но не могла проникнуть туда целиком, упираясь своим телом или инородным объектом (см. далее) в края отверстия, при этом совершая толкающие движения задними лапами в направлении отверстия.
Поведение животных в процессе эксперимента фиксировалось с помощью видеокамеры (Sony HDR-CX405) для off-line анализа данных.
Испытуемые были разделены на две группы:
• контрольную группу, в которой варьировались только характеристики внешней среды;
• экспериментальную группу, в которой варьировались как характеристики внешней среды, так и характеристики тела животных.
Процедура эксперимента. Перед началом эксперимента каждая жаба в течение 5 дней содержалась в отсеке № 2 для адаптации к экспериментальным условиям. При этом отверстия, разделяющие отсек № 1 и № 2, были закрыты.
В начале экспериментальной пробы жаба помещалась в центр отсека № 1. Она направлялась в отсек № 2 через одно из отверстий. Проба считалась завершенной, когда животное полностью покидало отсек № 1 через одно из отверстий. В ходе эксперимента проводилось 3 серии проб для контрольной группы и 4 серии проб для экспериментальной группы, каждая серия состояла из 30 проб (табл. 1). Каждая серия проводилось с интервалом в 24 часа, временной интервал между пробами внутри серии составлял 10 минут.
Описание экспериментальных серий контрольной группы
• Серия 1. Ставилась задача сформировать у жаб навык проникновения во второй отсек (рис. 3 А). Критерии сформированности навыка: значимое снижение количества неуспешных попыток проникновения в малые отверстия, сокращение времени решения задачи (аналогично в остальных сериях).
Таблица 1
Общая схема эксперимента
|
Серии |
Группы |
|
|
Экспериментальная |
Контрольная |
|
|
№ 1 |
Отверстия: № 1 — M; № 2 — S; № 3 — S. Границы тела — естественные |
|
|
№ 2 |
Отверстия: № 1 — M; № 2 — M; № 3 — L. Границы тела — измененные |
Отверстия: № 1 — S; № 2 — S; № 3 — M. Границы тела — естественные |
|
№ 3 |
Отверстия: № 1 — M; № 2 — L; № 3 — M. Границы тела — измененные |
Отверстия: №1 — S; №2 — M; №3 — S. Границы тела — естественные |
|
№ 4 |
Отверстия: № 1 — L; № 2 — M; № 3 — M. Границы тела — измененные |
|
• Серия 2. Схема расположения отверстий менялась таким образом, что отверстие M располагалось в другой части экспериментальной установки. Соответственно, животное не могло проникнуть в отсек № 2 ранее выученным маршрутом из-за изменения характеристик внешней среды.
• Серия 3. Схема расположения отверстий снова менялась таким образом, что отверстие M располагалось в другой части экспериментальной установки.
Гипотеза № 1 (о способности учитывать естественные границы собственного тела) может считаться подтвержденной в том случае, если в начале второй серии животное совершало достоверно меньше попыток проникновения в малые отверстия, нежели в начале первой серии.
Описание экспериментальных серий экспериментальной группы
• Серия 1. Первая экспериментальная серия была аналогична первой экспериментальной серии у контрольной группы животных. Также задачей являлось формирование навыка проникновения во второй отсек через отверстие № 1. Критерии сформированности навыка были такими же, как и в контрольной группе.
• Серия 2. Схема расположения отверстий менялась таким образом, что отверстия № 1 и № 2 делались размером M, а отверстие № 3 увеличивалось до L. Границы тела животных увеличивались. Соответственно, животное не могло проникнуть в отсек № 2 ранее выученным маршрутом не из-за изменения характеристик внешней среды, а из-за изменения характеристик его тела.
• Серия 3. Границы тела животного оставались увеличенными, а схема расположения отверстий снова менялась: проницаемое отверстие L теперь перемещалось в новое место. Животному было необходимо научиться проникать в отсек № 2 через отверстие L.
• Серия 4. Границы тела животного были увеличены, а схема расположения отверстий менялась.
Гипотеза № 2 (об учете увеличенных границ собственного тела) может считать подтвержденной в том случае, если в начале третьей серии животное совершало достоверно меньше попыток проникновения в средние отверстия, нежели в начале серии 2.
Статистическая обработка данных производилась с помощью критериев х2 и Теста Вилкоксона. Уровень значимости для обоих критериев составлял 0,05.

Рис. 3. А. Жаба, проникающая в отверстие M. Б. Жаба с увеличенными границами тела, совершающая неуспешную попытку проникновения в отверстие M
Результаты исследования
Контрольная группа
Серия 1. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 174 с (SD=42,1), в 30-й — 31 с (SD=12). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 149; в последних пяти пробах: успешных — 30, неуспешных — 2 (распределения достоверно отличаются друг от друга — X2=79,07; df=1; p<0,01).
Серия 2. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 87 с (SD=11,0), в 30-й — 32 с (SD=9,8). При этом на первой пробе 2-й серии испытуемые тратили меньше времени на решение задачи, нежели в первой пробе 1-й серии (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 4; в последних пяти пробах: успешных — 30, неуспешных — 4 (распределения не имеют достоверных отличий друг от друга — х2=0; df=1; p>0,05). При этом в серии 2 в первых пяти пробах количество неуспешных попыток проникновения достоверно ниже, нежели в первых пяти пробах серии 1 (х2=72,14; df=1; p<0,01).
Серия 3. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 80 с (SD=13,7), в 30-й — 31 с (SD=11,4). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 2; в последних пяти пробах: успешных — 30, неуспешных — 1 (распределения не имеют достоверных отличий друг от друга — х2=0,32; df=1; p>0,05).
Серия 1. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 165 с (SD=18,2), в 30-й — 32 с (SD=11). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 162; в последних пяти пробах: успешных — 30, неуспешных — 2 (распределения достоверно отличаются друг от друга — X2=85,37; df=1; p<0,01).
Серия 2. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 391 с (SD=74,5), в 30-й — 33 с (SD=8,8). При этом в первой пробе 2-й серии животные тратили больше времени на решение задачи, нежели в первой пробе 1-й серии (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 256; в последних пяти пробах: успешных — 30, неуспешных — 2 (распределения достоверно отличаются друг от друга — х2=130,33; df=1; p<0,01).
Серия 3. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 286 с (SD=58,1), в 30-й — 31 с (SD=10,1). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 274; в последних пяти пробах: успешных — 30, неуспешных — 4 (распределения достоверно отличаются друг от друга — х2=128,63; df=1; p<0,01). При этом в серии 3 в первых пяти пробах количество неуспешных попыток проникновения не имеет достоверных отличий в сравнении с количеством неуспешных попыток проникновения в первых пяти пробах серии 2 (х2=0,06; df=1; p>0,05).
Серия 4. От 1-й к 30-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (Тест Вилкоксона T=0; Z=2,20; n=6; p<0,05). Среднее время решения экспериментальной задачи в 1-й пробе составило 325 с (SD=93,6), в 30-й — 31 с (SD=11,4). В первых пяти пробах количество успешных попыток проникновения в отверстия составило 30, неуспешных попыток проникновения — 245; в последних пяти пробах: успешных — 30, неуспешных — 2 (распределения достоверно отличаются друг от друга — х2=125,10; df=1; p<0,01). При этом в серии 4 в первых пяти пробах количество неуспешных попыток проникновения не имеет достоверных отличий в сравнении с количеством неуспешных попыток проникновения в первых пяти пробах в серии 2 (х2=0,03; df=1; p>0,05).
Обсуждение результатов
Данные, полученные в серии 1, свидетельствуют о том, что к последним пяти пробам данной серии у испытуемых обеих выборок сформировался навык проникновения в отсек № 2 через отверстие M. Кроме того, на последних пяти пробах все испытуемые животные шли кратчайшим путем от места своего нахождения в начале пробы к нужному отверстию и проникали через него в отсек № 2.
Анализ данных показал, что жабы сформировали навык учета естественных границ собственного тела при взаимодействии с объектами внешней среды к концу серии № 1. Об этом свидетельствует тот факт, что в серии № 2 жабами из контрольной группы было совершено достоверно меньше неуспешных попыток проникновения в отверстия S, нежели
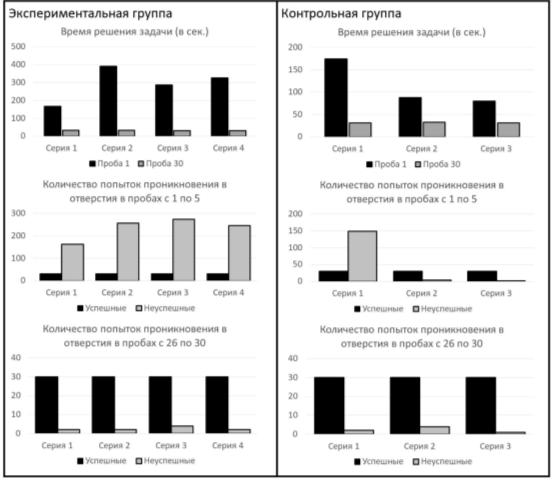
Рис. 4. Результаты эксперимента (пояснения см. в тексте)
в серии № 1. Следовательно, что в ходе серии № 1 жабы не только выучили схему расположения отверстий, но и начали воспринимать отверстие типа S непроницаемым для их тел. В серии № 2 животные сформировали навык проникновения в отсек № 2 новым маршрутом, практически не совершая неуспешных попыток проникновения в отверстия S. Аналогичные данные были получены в серии 3 для контрольной группы. Не было выявлено существенной динамики в соотношении успешных и неуспешных попыток проникновения в отверстия S между сериями 2 и 3 у контрольной группы.
Жабы из экспериментальной группы не смогли научиться учитывать экстренно увеличенные границы собственного тела, так как в первых пробах серий 3 и 4 количество неуспешных попыток проникновения в отверстия типа M возрастало до такого же значения, которое было зафиксировано в первых пяти пробах серии 2. К концу серий 3 и 4 количество неуспешных попыток проникновения в отверстия M снижалось и животные проникали в отсек № 2 сразу кратчайшим путем, но в начале серии 4 это количество неуспешных попыток снова возрастало. Это означает, что в серии 3 у жаб сформировался навык проникновения в отверстие, располагающееся в определенной части экспериментальной установки, однако пространственный навык по оценке отверстия L (но не M) как проницаемого вне зависимости от места его нахождения не сформировался.
Полученные данные объясняются тем, что для решения поставленной задачи испытуемым животным контрольной группы требовалось учитывать только изменения объектов окружающей среды (одну переменную). Испытуемым животным экспериментальной группы для решения их задачи в сериях 2—4 необходимо было учитывать как изменения внешних объектов, так и изменения характеристик собственного тела (2 переменные).
Кроме того, необходимо заметить, что у жаб вызывала большую трудность задача пространственной ориентации и определения размеров собственного тела, нежели задача с ориентированием и приспособлением к особенностям внешней среды. Соответственно, восприятие себя и формирование адекватного образа собственного тела для этих животных является более сложным процессом, нежели восприятие объектов внешней среды.
В более ранних исследованиях, проводившихся на тараканах Periplaneta americana (Хватов, 2011), а также на ряде других членистоногих, было установлено, что представители этого типа животных не формируют навыка учета ни естественных, ни увеличенных границ своего тела. Их навык решения задачи проявляется лишь в запоминании места расположения проницаемого отверстия (в каждой серии отдельно), но не в научении тому, что отверстия определенных размеров являются непроницаемыми для их тел инвариантно по отношению к месту их расположения.
Исследования, проводившиеся на лучистых полозах (Хватов и др., 2017) и синеязыких сцинках (Хватов и др., 2016 a) показали, что данные пресмыкающиеся способны научаться учитывать как естественные, так и увеличенные границы своего тела. Однако навык учета естественных границ складывался у них быстрее, нежели навык учета увеличенных границ.
При проведении аналогичного эксперимента на серых крысах (Хватов и др., 2016b) было установлено, что этим грызунам вовсе не требуется научаться учитывать естественные границы своих тел: даже наивные особи практически не совершали неуспешных попыток проникновения в отверстия типа S. Навык учета увеличенных границ тела складывался у них значительно быстрее, чем у пресмыкающихся — всего за 3—4 пробы в рамках одной серии. Более того, крысы оказались способны гибко менять свое поведение в зависимости от границ тела и расположения отверстий.
Таким образом, полученные нами данные согласуются с общими представлениями о наличии общей тенденции эволюции психики у животных, заключающейся в том, что отражение собственных характеристик (схемы тела) и характеристик внешних объектов в структуре единого образа мира все больше обособляются друг от друга. Данный тезис был сформулирован в работе И.А. Хватова и базируется на положениях онтологического подхода (Барабанщиков, 2002) и концепции воплощенного познания (Varela et al., 1991).
Выводы
На основании полученных данных необходимо заключить следующее. Жабы учитывают естественные границы собственного тела при взаимодействии с внешними объектами. При экстренном изменении границ тела жаб таким образом, что изменения препятствуют осуществлению животным ранее выученного поведения, животные не модифицируют схему своего тела и не способны развить навык учета этих объективных экстренных изменений при взаимодействии с объектами внешней среды, они лишь запоминают, где располагаются внешние объекты, с которыми не следует вступать в физический контакт.
Финансирование
Исследование поддержано грантом Совета по грантам при Президенте Российской федерации № МК-2766.2017.6.
Литература
-
Барабанщиков В.А. Восприятие и событие. СПб.: Алетейя, 2002. 512 с.
-
Ефанова Ю.Б., Матвеева Е.А., Бармашов А.E., Полозов В.М., Огурцов С.В. Ориентационное поведение серой жабы Bufo bufo L. в летний период // Флора и фауна западного Подмосковья. Студенческие самостоятельные работы, выполненные на Звенигородской биологической станции имени С.Н. Скадовского. Т. 5. М.: Изд-во Моск. ун-та имени М.В. Ломоносова, 2007. Т. 5. С. 81—95.
-
Кузьмин С.Л. Земноводные бывшего СССР. М.: Товарищество научных изданий КМК, 2012. 370 с.
-
Хватов И.А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011. Т. 4. № 1. C. 28—39.
-
Хватов И.А., Соколов А.Ю., Харитонов А.Н. Учет границ собственного тела сцинками Tiliqua gigas // Экспериментальная психология. 2016 a. Т. 9. № 3. С. 54—71. doi:10.17759/exppsy.2016090305
-
Хватов И.А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Схема собственного тела у грызунов (на примере крыс Rattus norvegicus) // Экспериментальная психология. 2016 b. Т. 9. № 1. С. 112—130. doi:10.17759/exppsy.2016090109
-
Хватов И.А., Соколов А.Ю., Харитонов А.Н. Сравнительный анализ восприятия физических характеристик собственного тела у змей lampropeltis triangulum campbelli и elaphe radiate // Экспериментальная психология. 2017. Т. 10. № 3. С. 126—136. doi:10.17759/exppsy.2017100309
-
Brattstrom B.H. Maze Learning in the Fire-Bellied Toad, Bombina orientalis // Journal of Herpetology. 1990. Vol. 24. № 1. P. 44—47.
-
Burghardt G.M. Environmental enrichment and cognitive complexity in reptiles and amphibians: Concepts, review, and implications for captive populations // Applied Animal Behaviour Science. 2013. Vol. 147(3, 4). P. 286—298.
-
Daneri M.F., Casanave E., Muzio R.N. Control of spatial orientation in terrestrial toads (Rhinella arenarum) // Journal of Comp. Psychol. 2011. Vol. 125(3). P. 296—307. doi: 10.1037/a0024242
-
Gallagher S., Cole J. Body Schema and Body Image in a Deafferented Subject // Journal of Mind and Behavior. 1995. Vol. 16. P. 369—390.
-
Johnson-Frey S. The neural bases of complex tool use in humans // Trends in Cognitive Sciences. 2004. Vol. 8(2). P. 71—78. doi:10.1016/j.tics.2003.12.002
-
Kundey S.M.A., Millar R., McPherson J., Gonzalez M., Fitz A., Allen C. Tiger salamanders’ (Ambystoma tigrinum) response learning and usage of visual cues // Anim Cogn. 2016. Vol. 19. P. 533—541.
-
Maravita A. Iriki A. Tools for the body (schema) // Trends in Cognitive Sciences. 2004. Vol. 8. № 2. P. 79—86.
-
Morasso P., Casadio M., Mohan V., Rea F., Zenzeri J. Revisiting the body-schema concept in the context of whole-body postural-focal dynamics // Front. Hum. Neurosci. 2015. 9:83. doi: 10.3389/fnhum.2015.00083
-
Sotelo M.I., Bingman V.P., Muzio R.N. Goal orientation by geometric and feature cues: spatial learning in the terrestrial toad Rhinella arenarum // Animal Cognition. 2015. Vol. 18(1). P. 315—323.
-
Varela F.J., Thompson E.T., Rosch E. The Embodied Mind: Cognitive Science and Human Experience. Cambridge, MA: The MIT Press, 1991.
Информация об авторах
Метрики
Просмотров
Всего: 1921
В прошлом месяце: 17
В текущем месяце: 12
Скачиваний
Всего: 788
В прошлом месяце: 6
В текущем месяце: 1