Экспериментальная психология
2014. Том 7. № 3. С. 137–144
ISSN: 2072-7593 / 2311-7036 (online)
Методика изучения схемы тела у мелких млекопитающих*
Аннотация
Общая информация
* Работа поддержана грантом Президента РФ № МК-5915.2014.6.
Ключевые слова: методика для изучения схемы тела, экспериментальная установка, мелкие млекопитающие, беспозвоночные
Рубрика издания: Методология психологических исследований
Тип материала: научная статья
Тематический сетевой сборник: 25 лет научных публикаций в журналах издательства МГППУ
Для цитаты: Хватов И.А., Соколов А.Ю., Харитонов А.Н., Куличенкова К.Н. Методика изучения схемы тела у мелких млекопитающих // Экспериментальная психология. 2014. Том 7. № 3. С. 137–144.
Полный текст
Одним из основных методов исследования специфики восприятия животными самих себя является тест с зеркалом (англ. “mirror-test”), в котором выявляется их способность опознавать собственное отражение. В классических экспериментах Г. Гэллапа (Gallup, 1970) шимпанзе под легким наркозом наносили пятнышки краски на одну из бровей и на противоположное ухо. После пробуждения животные не ощущали никаких физических последствий операций, проведенных с ними: они прикасались к этим частям тела не чаще, чем к остальным. Увидев же себя в зеркале, они начали активно ощупывать окрашенные места. Делался вывод, что шимпанзе помнили свой облик и замечали в нем изменения, а также понимали, что изображение в зеркале эквивалентно их собственному телу. На настоящий момент установлен факт наличия способности к самоузнаванию у понгид, макак-резусов, дельфинов, слонов, касаток и сорок (подробнее см.: Хватов, 2013).
Следует отметить, что данный метод подвергается существенной критике: во-первых, его трудно применять по отношению к животным, у которых зрительная модальность не является ведущей – например, к большинству млекопитающих (Bekoff, 2001), во-вторых, как животные, так и человек могут не проявлять интереса к отметинам на собственном теле (Asendorpf et al., 1996), в-третьих, дискуссионным остается вопрос о том, действительно ли самоузнавание в зеркале свидетельствует о самосознании (там же).
В рамках другого направления исследований изучается специфика схемы собственного тела у животных (Herman et al., 2001; Maravita, Iriki, 2004). Под схемой тела следует понимать систему двигательных навыков и способностей, позволяющих осуществлять различные движения, а также представление о физических характеристиках собственного тела (его границах, весе, взаиморасположении отдельных членов). В отличие от образа тела, схема тела является неосознаваемым феноменом (Gallagher, Cole, 1995). Существует точка зрения, что схема тела (принятие себя в расчет) является филогенетически наиболее ранней ступенью развития всех прочих представлений о себе, включая самосознание (Столин, 1983). В проведенных ранее исследованиях было показано, что животные способны включать в схему собственного тела внешние объекты (Maravita, Iriki, 2004).
На сегодняшний день в науке отсутствует отработанная методика, позволяющая ставить поведенческие эксперименты на млекопитающих с целью изучения особенностей их схемы тела. Однако за последние годы авторским коллективом данной статьи была разработана оригинальная методика, нацеленная на исследование специфики схемы тела беспозвоночных. Ниже будет приведено краткое описание данной методики.
В ходе эксперимента изучалось влияние изменения физических параметров тела животного на его поведение в различных экспериментальных установках. На начальном этапе эксперимента животное помещалось в экспериментальную установку (лабиринт или проблемную клетку) с целью формирования у него привыкания к данным условиям и осуществлению в них своей жизнедеятельности – в первую очередь кормления. Далее границы тела животного увеличивались экспериментатором с помощью крепления на него различных объектов таким образом, что они препятствовали осуществлению животным ранее сформированных двигательных навыков – проникновению в отверстия в перегородках, отделяющих один отсек лабиринта или проблемной клетки от другого. На других этапах эксперимента, наоборот, изменялись физические параметры среды (размеры отверстий в лабиринтах и проблемных клетках) при неизменности границ тела животного. Анализ данных о локомоторной активности животных, полученных в ходе этих экспериментов, позволил определить, способны ли животные учитывать физические параметры собственного тела при осуществлении поведения, а также способны ли они научаться менять свое поведение при изменении этих параметров и экстраполировать приобретенный опыт в новые ситуации. С помощью этой методики были получены данные о специфике психического отражения параметров собственного тела у тараканов, сверчков и моллюсков (Хватов, 2010, 2011; Хватов, Харитонов, 2012, 2013; Хватов, Харитонов, Соколов, 2013).
В настоящей работе мы предлагаем описание и апробацию методики и новой экспериментальной установки, ориентированной на исследование схемы тела млекопитающих, созданной на основе вышеизложенного опыта на беспозвоночных.
Метод
В качестве объекта исследования предполагается использование крыс и других мелких млекопитающих. Крысы и мыши являются модельными объектами в поведенческих исследованиях с применением различных типов лабиринтов (Barnes, 1979; Morris, 1984; Pellow et al., 1985; Takeda et al., 1998; Brown et al., 2009). Экологически и этологически обоснованной задачей для этих животных является проникновение в отверстия. Это обстоятельство используется в конструкциях некоторых лабиринтов (Barnes, 1979; Takeda et al., 1998). Для наших исследований мы создали экспериментальную установку, в которой животному было необходимо учитывать границы собственного тела при проникновении в различные отверстия для достижения приманки.
Оборудование. Для экспериментальных исследований сконструирован стеклянный лабиринт квадратной формы с пятью отсеками: центральным отсеком квадратной формы и четырьмя отсеками в форме равнобедренных трапеций, обрамляющими его. Обрамляющие отсеки сообщались между собой отверстиями в форме усеченного круга (диаметром 100 мм),
расположенными в боковых сторонах отсеков (рис. 1). Центральный отсек сообщался с каждым из четырех обрамляющих отсеков круглыми отверстиями (диаметром 80 мм). Диаметр отверстий центрального отсека мог уменьшаться с помощью дополнительных вставок до 40 мм (в это отверстие проникала только голова крысы) или 18 мм (в это отверстие проникала только передняя часть морды крысы). Таким образом, конструкция установки позволяла варьировать диаметр отверстий, через которые центральный отсек был доступен из боковых, что давало возможность планировать и чередовать различные экспериментальные серии.
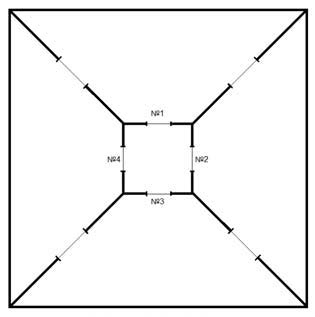
Рис. 1. Схема экспериментальной установки: номерами обозначены отверстия, через которые внешние отсеки сообщаются с центральным
Апробация
Испытуемые: 8 крыс Long-Evans (Rattus norvegicus), возраст 4 мес.
Фиксируемые показатели: время решения экспериментальной задачи (от момента помещения в экспериментальную установку до момента достижения приманки), количество проникновений/попыток проникновения в отверстия центрального отсека, время проникновений/попыток проникновения в отверстия центрального отсека.
Цель исследования: установить, способны ли крысы формировать навык получения приманки кратчайшим путем, о чем должно было свидетельствовать сокращение времени решения задачи, а также сокращение проникновений и попыток проникновения в различные типы отверстий.
Ход эксперимента. Эксперимент состоял из 20-и серий, проводившихся по две пробы ежедневно в 9 и 18 ч. В начале каждой пробы крыса запускалась во внешний отсек с отверстием № 1. В центральный отсек помещалась приманка (сыр). Расположение отверстий: № 1 – диаметром 40 мм, № 2, 3, 4 – диаметром 18 мм. Крысы обучались кормиться, просовывая голову в отверстие диаметром 40 мм (рис. 2).

Рис. 2. Крыса в экспериментальной установке
Результаты. У всех испытуемых от 1-й к 20-й пробе наблюдалось снижение времени решения задачи: на 1-й пробе у экспериментальной выборки среднее время решения составило 2187 с (SD=69,73), на 20-й пробе – 6 с (SD=1,83) (Wilcoxon Matched Pairs Test, T=0, p<0,01) (рис. 3). Также у всех крыс к 20-й пробе снижалось количество проникновений в различные отверстия: у экспериментальной выборки на 1-й пробе общее количество проникновений, считая то, которое привело к достижению приманки, составило 86, а на 20-й пробе – 8 (по одному на каждое животное); Wilcoxon Matched Pairs Test, T=0, p<0,01 (рис. 4).
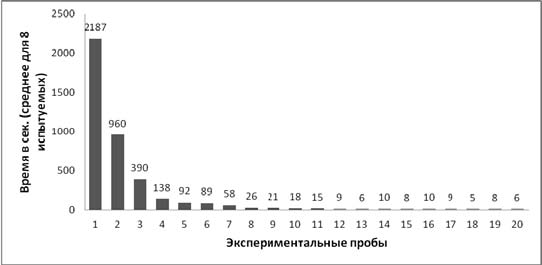
Рис. 3. Среднее время, затраченное на решение экспериментальной задачи
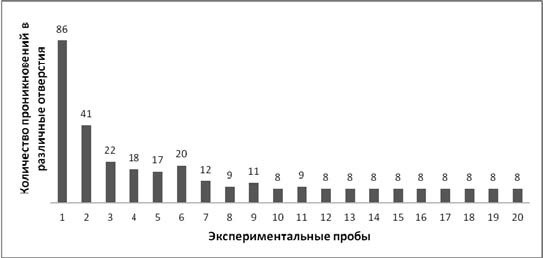
Рис. 4. Общее количество проникновений в различные отверстия
Обсуждение
Полученные данные свидетельствуют о том, что у крыс формируется навык достижения приманки кратчайшим путем через отверстие № 1. Соответственно, данная экспериментальная установка может быть использована для изучения особенностей научения у грызунов и, вероятно, других аналогичных по размеру млекопитающих. Для исследования схемы тела у животных можно ввести дополнительную экспериментальную переменную – варьирование границ тела животного с помощью закрепления на его теле различных легких конструкций.
Данные, полученные в ходе предыдущих исследований, проводившихся на беспозвоночных (Хватов, 2010, 2011; Хватов, Харитонов, 2012, 2013, Хватов, Харитонов, Соколов, 2013), позволяют заключить, что у ряда видов этой таксономической группы отсутствует схема тела в качестве целостной самостоятельной когнитивной структуры, данная схема складывается у них ситуативно в качестве элемента определенной перцептивной системы (целостного образа ситуации), формирующегося в процессе взаимодействия субъекта с конкретными условиями внешней среды. Если целостный образ ситуации у этих животных являлся перцептивным – в частности, в его структуре интегрируется визуальная и кинестетическая информация, – то отражение собственных характеристик, как компонент этого образа, остается сенсорным, поскольку формируется на основе одномодальной информации, преобладающей по силе и поступающей из определенного сегмента тела. На основе данных об основных тенденциях эволюции психики позвоночных и, в частности, млекопитающих (Филиппова, 2012; Хватов, 2012а, 2012b) можно выдвинуть гипотезу о том, что у позвоночных – возможно, уже у хладнокровных (пресмыкающихся) – возникает перцептивная схема тела, формирующаяся на основе интеграции сенсорных сигналов различных модальностей, поступающих от отдельных частей тела. Это позволяет животному антиципировать возможные последствия взаимодействия собственного тела с объектами в окружающем пространстве. Возможно, такая особенность в ходе прогрессивной эволюции привела к формированию обобщенного психического образа собственного тела у млекопитающих и птиц, и именно это качественно отличает психику позвоночных от психики беспозвоночных, схема тела которых представляет собой совокупность сенсорных сигналов, не связанных (или очень слабо связанных) между собой.
На наш взгляд, предложенная в в настоящей статье методика может быть использована в ходе сравнительно-психологических экспериментов, нацеленных, в частности, на проверку вышеизложенной гипотезы.
Литература
- Столин В.В. Самосознание личности. М.: Издательство Московского Университета, 1983. 284 с.
- Филиппова Г.Г. Зоопсихология и сравнительная психология: учеб. пособие для студентов вузов. 6-е изд., перераб. М.: Академия, 2012. 543 с.
- Хватов И.А. Особенности самоотражения у животных на разных стадиях филогенеза. Дисс. канд. психол. наук. М., 2010.
- Хватов И.А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011. Т. 4. № 1. C. 28–39.
- Хватов И.А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 1 // Психологические исследования. 2012а. № 1 (21). С. 1. URL: http://psystudy.ru/index.php/num/2012n1-21/622-khvatov21.html (дата обращения: 22.09.2014). 0421200116/0001.
- Хватов И.А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 2 // Психологические исследования. 2012b. № 2 (22). С. 12. URL: http://psystudy.ru/index.php/num/2012n2-22/648-khvatov22.html (дата обращения: 22.09.2014). 0421200116/0024.
- Хватов И.А. Проблема интеллекта животных в контексте структурно-интегративного и дифференционно-интеграционного подходов // Психологические исследования. 2013. Т. 6, № 28. С. 1. URL: http://psystudy.ru/index.php/num/2013v6n28/788-khvatov28.html (дата обращения: 28.07.2014).
- Хватов И.А., Харитонов А.Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. Т. 5. № 3. C. 96-107.
- Хватов И.А., Харитонов А.Н. Модификация плана развертки собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica // Экспериментальная психология. 2013. Т. 6. № 2. С. 101-114.
- Хватов И.А., Харитонов А.Н., Соколов А.Ю. Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Gryllus assimilis // Экспериментальная психология. 2013. Т. 6. № 4. С. 79-95.
- Asendorpf J. B., Warkentin V., Baudonniere P.-M. Self-Awareness and Other-Awareness II: Mirror Self-Recognition, Social Contingency Awareness, and Synchronic Imitation // Developmental Psychology. 1996. V. 32. № 2. P. 313-321.
- Barnes C.A. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat // J. Comp. Physiol. Psychol. 1979. V. 93. № 1. P. 74-104.
- Bekoff M. Observations of scent-marking and discriminating self from others by a domestic dog (Canis familiaris): tales of displaced yellow snow // Behavioural Processes. 2001. V. 55. № 2. P. 75-79.
- Brown M.F., Prince T.N., Doyle K.E. Social effects on spatial choice in the radial arm maze // Learning & Behavior. 2009. V. 37. № 3. P. 269-280.
- Gallagher S., Cole J. Body Schema and Body Image in a Deafferented Subject // Journal of Mind and Behavior. 1995. Vol. 16. P. 369-390.
- Gallup G. G. Jr. Chimpanzees: Self recognition // Science. 1970. V. 167. № 3914. P. 86—87.
- Herman L.M., Matus D.S., Herman E.Y.K., Ivancic M., Pack A.A. The bottlenosed dolphin’s (Tursiops truncatus) understanding of gestures as symbolic representations of its body parts // Animal Learning & Behavior. 2001. V. 29. № 3. P. 250-264.
- Maravita A., Iriki A. Tools for the body (schema) // Trends in Cognitive Sciences. 2004. V. 8. № 2. P. 79–86.
- Morris R. Developments of a water-maze procedure for studying spatial learning in the rat // Journal of neuroscience methods. 1984.V. 11. № 1. P. 47–60.
- Pellow S., Chopin P., File S.E., Briley M. Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat // J. Neurosci. Methods. 1985. V. 14. P. 149–167.
- Takeda H., Tsuji. M., Matsumiya T. Changes in head-dipping behavior in the hole-board test reflect the anxiogenic and/or anxiolytic state in mice // European Journal of Pharmacology. 1998. V. 350. № 1. P. 21–29.
Информация об авторах
Метрики
Просмотров
Всего: 3022
В прошлом месяце: 16
В текущем месяце: 23
Скачиваний
Всего: 747
В прошлом месяце: 1
В текущем месяце: 3