Экспериментальная психология
2015. Том 8. № 2. С. 119–138
doi:10.17759/exppsy.2015080209
ISSN: 2072-7593 / 2311-7036 (online)
Схема собственного тела у змей Lampropeltis triangulum campbelli*
Аннотация
Общая информация
* Работа поддержана грантом РГНФ № 14-06-00669, тип проекта – а.
Ключевые слова: самоотражение, схема тела, змеи, пресмыкающиеся, членистоногие, крысы, филогенез психики
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2015080209
Для цитаты: Хватов И.А., Соколов А.Ю., Харитонов А.Н. Схема собственного тела у змей Lampropeltis triangulum campbelli // Экспериментальная психология. 2015. Том 8. № 2. С. 119–138. DOI: 10.17759/exppsy.2015080209
Полный текст
Введение
При ориентации в окружающем пространстве для осуществления локомоции и манипуляции животным необходимо учитывать физические характеристики собственного тела (границы, объем, массу) и соотносить их с физическими характеристиками внешних объектов. Иначе говоря, животным необходимо «принимать себя в расчет» (Столин, 1983). Человек и другие приматы осуществляют этот процесс преимущественно с опорой на зрение – для нас зрительная система является ведущей при построении когнитивной репрезентации внешнего мира и самих себя. Поэтому одним из существенных критериев сознания считается способность узнавать собственное отражение в зеркале. Этот феномен, начиная с классических экспериментов Г. Гэллапа (Gallup, 1970), активно исследуется на детях и на детенышах других млекопитающих. На сегодняшний день установлен факт наличия способности к самоузнаванию у понгид, макак-резусов, слонов, дельфинов, касаток и сорок (подробнее см.: Хватов, 2013). Следует отметить, что данный метод подвергается существенной критике: во-первых, его трудно применять по отношению к животным, у которых зрительная модальность не является ведущей (например, к большинству млекопитающих; см.: Bekoff, 2001), во-вторых, как животные, так и человек могут и не проявлять интереса к отметкам, наносимым на тело, что используется в таких исследованиях как один из основных экспериментальных приемов (Asendorpf et al., 1996), в-третьих, дискуссионным остается вопрос о том, действительно ли самоузнавание в зеркале свидетельствует о самосознании (там же).
В рамках близкого направления исследований изучается «схема тела» человека и других животных (Herman et al., 2001; Maravita, Iriki, 2004). Схема тела – это совокупность двигательных навыков и способностей, позволяющих осуществлять различные движения, а также представление о физических характеристиках собственного тела (его границах, весе, взаиморасположении отдельных членов). В отличие от образа тела, схема тела является неосознаваемым феноменом (Gallagher, Cole, 1995). Существует точка зрения, что схема тела («принятие себя в расчет») является филогенетически наиболее ранней ступенью развития всех прочих представлений о себе, включая самосознание (Столин, 1983). Ранее было показано, что животные способны включать в схему собственного тела внешние объекты (Maravita, Iriki, 2004).
Более 10 лет одним из авторов данной статьи разрабатывается концепция самоотражения животных и человека (Хватов, 2010; 2014), базирующаяся на онтологическом подходе (Барабанщиков, 2002). В рамках данной концепции схема тела рассматривается как один из продуктов процесса самоотражения. Самоотражение определяется как процесс и результат отражения субъектом своей внутренней объективной реальности: характеристик своего организма, а также, если речь идет о человеке, своих свойств в качестве участника социальных процессов. Самоотражение рассматривается системно, в контексте своей взаимосвязи с отражением внешней среды, как двух элементов единого психического отражения индивидом акта своего события с окружающей объективной реальностью.
Авторским коллективом настоящей статьи было дано целостное описание процесса развития самоотражения в ходе филогенеза животных (Хватов, 2010; 2014), а также в рамках филогенеза отдельных крупных таксонов (Хватов и др., 2013b; 2014). Также был проведен ряд экспериментальных исследований особенностей процесса самоотражения и схемы тела у отдельных видов животных (Хватов, 2010; 2011; Хватов, Харитонов, 2012; 2013; Хватов и др., 2013a; 2014a; 2014b). В частности, нами была разработана и прошла успешную апробацию оригинальная экспериментальная методика изучения особенностей схемы тела у малоизученной в этом отношении группы – змей (Хватов и др., 2015). В рамках настоящей работы с помощью этой методики было осуществлено полномасштабное исследование схемы собственного тела у королевских змей (Lampropeltis triangulum campbelli).
Гипотеза исследования. Мы полагаем, что при изменении границ тела змей таким образом, что данные изменения будут препятствовать осуществлению животным ранее выученного поведения, данные животные модифицируют схему своего тела, что будет выражаться в том, что они модифицируют свое прежнее поведение с учетом изменений собственного тела.
Цель исследования: изучить влияние объективных изменений границ тела змей на специфику их схемы собственного тела.
Методика исследования
Испытуемые животные: 18 змей: 6 самцов и 10 самок королевской змеи Lampropeltis
triangulum campbelli. Возраст змей – около 6 месяцев. Змеи подбирались по
размеру таким образом, что максимальный диаметр сечения тела у них составлял
около 12–13 мм, поскольку этого требовали условия экспериментальной
методики.
Для данного вида змей является экологичным роющее поведение, они предпочитают находиться в темном и влажном окружении.
Животные были разделены на 2 группы: экспериментальную (8
особей) и контрольную (8 особей).
Оборудование. Экспериментальная установка представляет собой стеклянный ящик с
двумя отсеками (рис. 1).
Отсек А – пусковая камера 310×390 мм. Пол покрыт шпатовой и мраморной окатанной галькой белого цвета (для удобства различения змеи на субстрате при видеосъемке).
Отсек Б – «влажная» камера (укрытие) 250×390 мм. В качестве субстрата использовалась сосновая кора-мульча, сфагнум, крошка кокосовой коры. Субстрат увлажнялся исходно и затем дополнительно каждые 2 дня. В этом отсеке располагалась поилка. Стены и потолок отсека с внешней стороны были окрашены черной светонепроницаемой краской, благодаря чему в нем постоянно поддерживался низкий уровень освещенности.
Камеры разделены между собой стеклянной перегородкой, также
окрашенной в черный цвет. В перегородке имеются три круглых отверстия диаметром
70 мм, расположенных на уровне 5 мм от пола. Диаметр отверстий можно
варьировать с помощью дополнительных вставок.
В эксперименте использовались отверстия трех диаметров:
- большое отверстие (D=70 мм, без вставки) – в данное отверстие змея могла проникнуть без затруднений даже после проглатывания пищи (далее – L);
- среднее отверстие (вставка с отверстием D=14 мм) – в данное отверстие змея проникала без затруднений за исключением случаев, когда ее тело было увеличено после заглатывания пищи (далее – M);
- малое отверстие (вставка с отверстием D=8 мм) – в данное отверстие змея проникнуть не могла (далее – S).
Температурный режим. Эксперимент проводился при стандартных условиях обогрева. Область обогрева была оборудована с помощью термокабеля, пропущенного под террариумом, ее площадь составляла около 1/3 задней части отсека Б, температура 30–32°C. Температура в остальной части террариума составляла 25–26°C.

Рис. 1. Слева – Схема экспериментальной установки: внизу отсек А (светлый), вверху отсек Б (темный); номерами обозначены отверстия (см. в тексте). Справа – змея, совершающая попытку проникновения во «влажную» камеру экспериментальной установки
Процедура эксперимента. Перед началом эксперимента змея в течение семи дней содержалась в экспериментальной установке с тремя отверстиями типа M.
В начале каждой пробы змея помещалась в центр отсека А. Нахождение на жестком субстрате в светлом помещении, лишенном каких-либо укрытий, являлось отрицательной стимуляцией для животного, и оно стремилось покинуть отсек А и проникнуть в отсек Б («влажную» камеру) через одно из трех отверстий (рис. 1). Время пробы не ограничивалось; проба считалась завершенной, когда змея осуществляла успешную попытку проникновения во «влажную» камеру через отверстие L или M, т. е. когда ее тело полностью покидало пусковую камеру. Между экспериментальными пробами внутри серии выдерживался временной интервал в 15 минут.
Все экспериментальные серии проводились последовательно с каждой змеей из обеих групп по отдельности.
Эксперимент состоял из трех серий, каждая по 20 проб (табл. 1).
Схема организации эксперимента
Таблица 1
|
|
Группы |
|
|
Серии |
Экспериментальная |
Контрольная |
|
№ 1 |
Отверстия: №1 – M; №2 – S; №3 – S. Границы тела – естественные | |
|
№ 2 |
Отверстия: №1 – M; №2 – S; №3 – L. Границы тела – измененные |
Отверстия: №1 – S; №2 – S; №3 – M. Границы тела – естественные |
|
№ 3 |
Отверстия: №1 – L; №2 – S; №3 – M. Границы тела – измененные |
Отверстия: №1 – M; №2 – S; №3 – S. Границы тела – естественные |
Серия № 1. Ставилась задача сформировать у змей обеих выборок навык проникать в отсек Б через отверстие № 1 (лишь оно имело достаточный диаметр для проникновения через него тела животного). О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества попыток проникновения в отверстия № 2 и № 3.
Серия № 2. Спустя 24 часа после окончания серии № 1 у змей экспериментальной выборки увеличивались границы тела, благодаря чему они были неспособны проникнуть целиком в отверстие типа M, и, соответственно, могли покинуть пусковую камеру лишь через отверстие типа L. Границы тела животного увеличивались благодаря скармливанию змее кормового объекта – мыши субадультного возраста (длина тела – около 5 см). Кормление осуществлялось в отсеке № 2, после кормления до начала экспериментальной серии был выдержан временной промежуток в 12 часов. Проглоченный кормовой объект располагался в желудке змеи (начало второй половины длины тела) благодаря чему сечение тела увеличивалось, делая невозможным проникновение в отверстия типа M. Ставилась задача выявить, смогут ли змеи при условии увеличения границ их тела до такого размера, что решение прежней задачи ранее выученным способом (проникновение в отсек № 2 через отверстие № 1) окажется невозможным, модифицировать собственное поведение для достижения желаемого результата, т.е. найти новый путь в отсек Б. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества попыток проникновения в отверстия № 1 и № 2.
В контрольной выборке границы тела змей оставались неизменными, однако отверстие № 1, через которое в предыдущей серии у них был сформирован навык покидания пускового отсека, делалось непроницаемым (S), проницаемым (M) делалось отверстие № 3. Ставилась задача выявить, смогут ли змеи при условии неизменности физических параметров тела, но изменении условий внешней среды, найти новый путь в отсек Б. Полученные данные (по количеству проникновений в отверстия и времени решения экспериментальной задачи) сопоставлялись с данными, полученными на змеях экспериментальной выборки в этой же серии, чтобы установить, по-разному ли влияет на поведение змей ситуация невозможности реализации ранее сформированного навыка, создаваемая изменением параметров тела в экспериментальной группе и изменением параметров внешней среды в контрольной группе.
Серия № 3. Проводилась спустя 3 часа после окончания серии № 2 и при условии, что к концу серии № 2 у испытуемых обеих выборок удалось сформировать новый навык проникновения в отсек Б через отверстие № 3.
В экспериментальной группе ставилась задача установить, как быстро и после какого количества ошибок (попыток проникновения в слишком маленькие для их тел отверстия) змеи смогут сформировать новый навык проникновения в отсек Б через отверстие № 1 при условии, что границы тела останутся увеличенными, но размер отверстия № 3 уменьшится до M, т.е. станет непроницаемым для их тела. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества попыток проникновения в отверстия № 2 и № 3.
В контрольной выборке границы тела оставались неизменными, расположение отверстий было идентично тому, что было в серии № 1. Ставилась задача, смогут ли змеи сформировать новый навык проникновения в отсек Б при условии неизменности границ их тела. Полученные данные (по количеству проникновений в отверстия и времени решения экспериментальной задачи) сопоставлялись с данными, полученными на змеях экспериментальной выборки в этой же серии.
Гипотеза исследования могла считаться подтвержденной в том случае, если испытуемые экспериментальной выборки в серии № 3 совершали достоверно меньше попыток проникновения в отверстия типа M, чем в серии № 2 в аналогичные отверстия. Это свидетельствовало бы о том, что при формировании навыка змеи не только выучивали новое расположение отверстий, но и начинали учитывать увеличение границ собственного тела, т.е. модифицировали собственную схему тела. Дополнительным основанием подтверждения гипотезы можно было считать более быстрое формирование навыка в серии № 3, нежели в серии № 2, у испытуемых экспериментальной выборки.
Аппаратура. Поведение животных в процессе эксперимента фиксировалось с помощью видеокамеры (Sony HDR-CX405), для фиксации временных интервалов использовался таймер видеозаписи данной камеры.
Регистрируемые показатели.
-
Время (в секундах) решения экспериментальной задачи в каждой пробе от момента помещения змеи в пусковую камеру до момента полного проникновения животного во влажную камеру через одно из отверстий;
-
Количество попыток проникновений (успешных и неуспешных) в различные типы отверстий отдельно для каждого отверстия в каждой пробе и отдельно для каждой змеи в каждой пробе. За одну попытку проникновения считались ситуации, когда змея погружала голову в отверстие типа M или L хотя бы на несколько миллиметров, а также ситуации, в которых змея прижималась мордой к отверстию типа S, совершая в направлении него ундуляционные (извивающиеся) движения телом.
Особое внимание было уделено неуспешным попыткам проникновения в отверстия типа M, совершенным змеями экспериментальной группы с увеличенными границами тела в сериях № 2 и № 3, что объяснялось их существенным разнообразием. В ходе предварительных пилотажных исследований было установлено, что эти попытки имеют значительный разброс по продолжительности (от 52 с до 1 с), а также имеют разнообразные поведенческие проявления.
-
В одних случаях змеи проникали в отверстия глубоко (часто до области желудка с проглоченным кормовым объектом), при этом либо совершая интенсивные ундуляционные движения в направлении отверстия, либо совершая возвратно-поступательные движения частью тела, находящейся в отверстии, но при этом не покидая его целиком. Такие проникновения, как правило, занимали длительные временные промежутки. Такие попытки в дальнейшем будут называться длительными попытками проникновения. Обратим внимание, что сходные длительные попытки проникновения ранее наблюдались у крыс с увеличенными границами головы, когда эти животные, стремясь достать приманку, максимально глубоко проникали в отверстия, совершая при этом буксирующие движения лапами в направлении отверстия, а также могли осуществлять возвратно-поступательные движения головой (Хватов и др., 2014b).
-
В других случаях змеи проникали в отверстие только головой или частью головы, не достигая области с проглоченным кормовым объектом, и завершали попытку. Эти проникновения, как правило, занимали короткие временные промежутки. Такие попытки в дальнейшем будут называться краткими попытками проникновения. Также обратим внимание, что сходные попытки проникновения обнаруживались у крыс с увеличенными границами головы, в ходе которых животные проникали в отверстие лишь передней частью морды, не совершая буксирующих движений лапами (у крыс такие попытки проникновения были названы ориентировочными проникновениями в отверстия). Соответственно, для дальнейшего разделения попыток проникновения на длительные и краткие фиксировались следующие показатели неуспешных попыток проникновения в отверстия типа M, совершенных испытуемыми экспериментальной выборки в каждой пробе серий № 2 и № 3.
-
Длительность каждой попытки проникновения (в секундах), считавшаяся от момента, когда змея погружала голову в отверстие хотя бы на несколько миллиметров, до момента, когда змея полностью вынимала голову из отверстия.
-
Глубина проникновения в отверстие. Для большей объективности использовалась дихотомическая шкала на основе анатомических характеристик животного: проникает ли в отверстие только голова животного, что было характерно для кратких попыток, или же змея проникала в отверстие глубже, что было характерно для длительных попыток.
На основе этих критериев все вышеописанные попытки были дифференцированы на длительные и краткие и отдельно зарегистрированы в каждой пробе серий № 2 и 3 у испытуемых экспериментальной группы.
Также в ходе пилотажных исследований было установлено, что в рамках одной пробы частота встречаемости длительных и кратких попыток проникновения оказывалась не одинаковой. Максимально за одну пробу змея с увеличенными границами тела могла совершить до 10 неудачных попыток проникновения в отверстия типа M. При этом длительные попытки проникновения чаще встречались в начале пробы, краткие – в конце. Длительные попытки проникновения занимают больше времени и требуют больших физических усилий. Мы предположили, что у змей с увеличенными границами тела о формировании навыка решения экспериментальной задачи свидетельствуют не только количественные данные (снижение времени, затрачиваемого на решение задачи и снижение количества ошибочных попыток проникновения), но и изменение качества ошибочных попыток проникновения в отверстия типа M – уменьшение доли длительных попыток проникновения и возрастание доли кратких в рамках каждой отдельной пробы.
Соответственно, у животных экспериментальной группы в сериях № 2 и 3 фиксировалась последовательность распределения длительных и кратких попыток проникновения в отверстия типа M в рамках каждой пробы в отдельности.
Переменные. Независимые переменные:
- границы тела испытуемых змей: естественные или увеличенные;
- диаметр отверстий в перегородке, разделяющей отсеки А и Б экспериментальной установки.
Зависимые переменные:
- время решения экспериментальной задачи в каждой пробе;
-
количество неуспешных попыток проникновения в отверстия типов S и M (в том числе, количество длительных и кратких попыток проникновения у испытуемых экспериментальной группы в сериях № 2 и № 3).
Побочные переменные:
-
пол испытуемых змей. Отметим, что в ходе эксперимента не было выявлено значимых различий между самцами и самками ни по одному из фиксируемых показателей.
Статистические критерии и статистические гипотезы.
-
Для выявления формирования навыка по показателю времени решения задачи использовался T-критерий Вилкоксона (n=20; p<0,01). Сопоставлялись временные показатели решения экспериментальной задачи испытуемыми конкретной группы на 1-й и 20-й пробах. В качестве типичного сдвига бралось сокращение времени.
-
Для выявления формирования навыка по критерию количества проникновений в отверстия использовался критерий Ȥ2 (df=2; p<0,01). Сопоставлялись распределения попыток проникновения в каждое из трех отверстий за первые 5 проб и последние 5 – в конкретной серии у конкретной группы. H : распределения не различаются. H : распределения различаются между собой при том, что на последних 5 пробах змеи чаще совершают попытки проникновения в проницаемые отверстия и реже – в непроницаемые.
-
Для сопоставления экспериментальной и контрольной выборок по показателю времени решения задачи использовался U-критерий Манна–Уитни (n =8; n =8; p<0,01). Cопоставлялись общее время решения экспериментальной задачи за 20 проб конкретной серии у животных экспериментальной и контрольной выборок. H : испытуемые ни одной из выборок не затратили больше времени на решение экспериментальной задачи в конкретной серии. H : испытуемые одной из выборок затратили больше времени на решение экспериментальной задачи, чем испытуемые другой выборки в конкретной серии.
-
Для сопоставления экспериментальной и контрольной выборок по показателю количества попыток проникновения в различные отверстия использовался критерий Ȥ2 (df=2; p<0,01). Сопоставлялись распределения попыток проникновения в каждое из трех отверстий за все пробы конкретной серии у животных экспериментальной и контрольной выборок. H : распределения не различаются. H : распределение экспериментальной выборки отличается от распределения контрольной выборки.
-
Для сопоставления динамики решения задач в разных сериях по показателю времени у испытуемых конкретной группы использовался T-критерий Вилкоксона (n=20; p<0,01). Сопоставлялось среднее время решения задачи (восьмью змеями конкретной выборки) в каждой пробе в двух экспериментальных сериях. H : между двумя сериями в конкретной группе отсутствуют значимые сдвиги по времени решения задачи. H : между двумя сериями в конкретной группе имеются значимые сдвиги по времени решения задачи.
-
Для сопоставления долей неуспешных попыток проникновения в отверстия типа M в общем числе попыток проникновения, совершенных испытуемыми экспериментальной выборки в сериях № 2 и № 3 использовался критерий Ȥ2 (df=2; p<0,01). Сопоставлялись распределения попыток проникновения в отверстия типов S, M и L (безотносительно их положения в экспериментальной установке) за все пробы у испытуемых экспериментальной выборки в сериях № 2 и № 3. H : распределения не различаются. H : распределение в серии № 2 отличается от распределения в серии № 3.
-
Для разделения всех неудачных попыток проникновения в отверстия типа M, совершенных испытуемыми экспериментальной выборки в сериях № 2 и № 3, на длительные и краткие был применен кластерный анализ по методу k-средних с выделением двух кластеров. Переменные для кластеризации: длительность попытки проникновения (в секундах), глубина проникновения в отверстие; была проведена предварительная стандартизация данных.
-
Полученные в результате кластерного анализа длительные и краткие попытки проникновения были разбиты на 10 разрядов в зависимости от того, какими по счету они являлись в рамках последовательности проникновений, совершенных в каждой отдельной пробе серий № 2 и № 3. Эмпирические распределения длительных и кратких попыток проникновения были сопоставлены с помощью критерия Ȥ2 (df=9; p<0,01). H : распределения не различаются. H : эмпирическое распределение порядковых номеров длительных проникновений отличается от эмпирического распределения порядковых номеров кратких проникновений.
-
Для сопоставления распределений длительных и кратких попыток проникновения, совершенных испытуемыми экспериментальной группы в сериях № 2 и № 3 использовался критерий Ȥ2 (df=1; p<0,01). H : распределения не различаются. H : эмпирическое распределение в серии № 2 отличается от эмпирического распределения в серии № 3.
Все расчеты (за исключением кластерного анализа) проводились для испытуемых каждой группы в каждой из трех экспериментальных серий. Математический анализ осуществлялся с помощью программы Statistica 8.
Результаты
Серия № 1. У испытуемых обеих групп от 1-й к 20-й пробе
наблюдалось снижение времени решения экспериментальной задачи (рис. 2) (в
экспериментальной выборке: Тест Вилкоксона T=0; Z=2,52; n=8; p<0,01, в
контрольной выборке: T=0; Z=2,52; n=8; p<0,01). Среднее время решения
экспериментальной задачи на 1-й пробе в экспериментальной группе составило 234
с (SD=17,8), в контрольной группе – 229 с (SD=16,9). Среднее время решения
экспериментальной задачи на 20-й пробе в экспериментальной группе составило 12
с (SD=1,7), в контрольной группе – 11 с (SD=1,6). По показателям времени
решения экспериментальной задачи испытуемые экспериментальной и контрольной
выборок не отличались друг от друга (Тест Манна–Уитни U=16,5; n =8; n =8;
p>0,05).
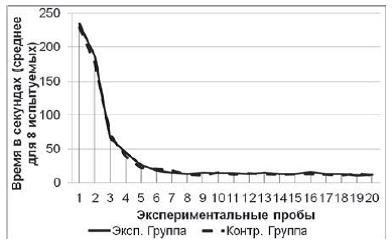
Рис. 2. Кривые научения контрольной и экспериментальной групп в серии № 1. Показано среднее время решения задачи в каждой пробе для восьми испытуемых в каждой группе. К концу пробы испытуемые обучались, так как для решения задачи им требовалось значительно меньше времени, нежели в начале серии (Тест Вилкоксона; p<0,01)
В экспериментальной и контрольной группах к концу серии змеи стали чаще совершать попытки проникновения в отверстия № 1 и реже – в отверстия № 2 и № 3 (рис. 3). В экспериментальной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 21%, на последних 5 пробах – 87%; Ȥ2=74,7; df=2; p<0,01. В контрольной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 23%, на последних 5 пробах – 91%; Ȥ2=69,7; df=2; p<0,01.
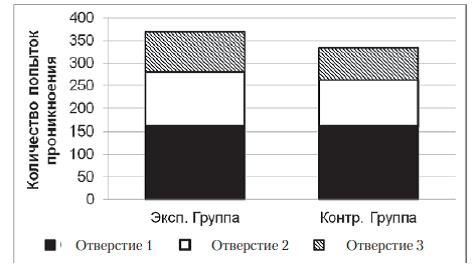
Рис. 3. Распределения общего количества проникновений в различные отверстия в контроль- ной и экспериментальной группах в серии № 1. Распределения не имеют достоверных отличий друг от друга (Ȥ2=1,7; df=2; p>0,05)
Серия № 2. У испытуемых обеих групп от 1-й к 20-й пробе
наблюдалось снижение времени решения экспериментальной задачи (рис. 4) (в
экспериментальной выборке: T=0; Z=2,52; n=8; p<0,01, в контрольной выборке:
T=0; Z=2,52; n=8; p<0,01). Среднее время решения экспериментальной задачи на
1-й пробе в экспериментальной группе составило 420 с (SD=25,6), в контрольной
группе – 219 с (SD=15,2). Среднее время решения экспериментальной задачи на
20-й пробе в экспериментальной группе составило 21 с (SD=7,4), в контрольной
группе – 13 с (SD=1,5). При этом на решение экспериментальной задачи испытуемые
экспериментальной группы затратили больше времени, нежели испытуемые
контрольной группы (U=0; n =8; n =8; p<0,01). Также змеи экспериментальной
выборки в серии № 2 тратили больше времени на решение экспериментальной задачи,
нежели в серии № 1 (T=7,5; Z=3,64; n=20; p<0,01), у испытуемых контрольной
выборки таких различий между сериями не было выявлено (T=102; Z=0,11; n=20;
p>0,05).
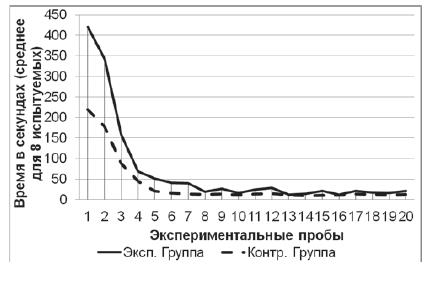
Рис. 4. Кривые научения контрольной и экспериментальной групп в серии № 2. Показано среднее время решения задачи в каждой пробе для восьми испытуемых в каждой группе. К концу пробы испытуемые обучались, так как для решения задачи им требовалось значительно меньше времени, нежели в начале серии (Тест Вилкоксона; p<0,01)
В экспериментальной и контрольной группах к концу серии змеи стали чаще совершать попытки проникновения в отверстия № 3 и реже в отверстия № 1 и № 2. В экспериментальной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 14%, на последних 5 пробах – 87%; Ȥ2=112,7; df=2; p<0,01. В контрольной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 23%, на последних 5 пробах – 95%; Ȥ2=76,5; df=2; p<0,01 (рис. 5). При этом распределение попыток проникновения в три отверстия во всех пробах серии № 2 у испытуемых экспериментальной группы отличается от аналогичного распределения у испытуемых контрольной группы (Ȥ2=18,5; df=2; p<0,01). В частности, доля неуспешных попыток проникновения в отверстие № 1 (через это отверстие испытуемые покидали пусковую камеру в предыдущей серии) за все 20 проб серии № 2 у испытуемых экспериментальной выборки составляет 43%, а у испытуемых контрольной выборки – 29%. Змеи экспериментальной выборки чаще совершали попытки проникновения в отверстие № 1.
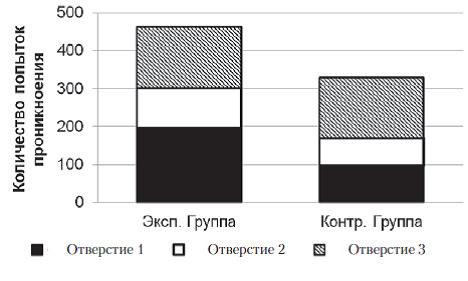
Рис. 5. Распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 2. Распределения достоверно отличаются друг от друга (Ȥ2=18,5; df=2; p<0,01)
Серия № 3. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 6) (в экспериментальной выборке: T=0; Z=2,52; n=8; p<0,01, в контрольной выборке: T=0; Z=2,52; n=8; p<0,01). Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 402 с (SD=25,7), в контрольной группе – 215 с (SD=17,4). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 16 с (SD=2,0), в контрольной группе – 12 с (SD=2,0). Как и в предыдущей серии, на решение экспериментальной задачи испытуемые экспериментальной группы затратили больше времени, нежели испытуемые контрольной группы (U=0; n1=8; n2=8; p<0,01). Однако временные интервалы, затраченные змеями экспериментальной выборки на решение экспериментальной задачи в серии № 3, не изменились в сравнении с интервалами, затраченными этими же змеями в серии № 2 (T=104; Z=0,04; n=20; p>0,05), аналогичная ситуация обнаруживается и в контрольной выборке (T=62; Z=0,31; n=20; p>0,05).
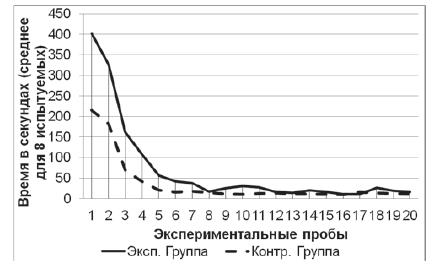
Рис. 6. Кривые научения контрольной и экспериментальной групп в серии № 3. Показано среднее время решения задачи в каждой пробе для восьми испытуемых в каждой группе. К концу пробы испытуемые обучались, так как для решения задачи им требовалось значительно меньше времени, нежели в начале серии (Тест Вилкоксона; p<0,01)
В экспериментальной и контрольной группах к концу серии змеи стали чаще совершать попытки проникновения в отверстие № 1 и реже – в отверстия № 2 и № 3. В экспериментальной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 15%, на последних 5 пробах – 82%; Ȥ2=101,1; df=2; p<0,01. В контрольной выборке на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 21%, на последних 5 пробах – 87%; Ȥ2=68,5; df=2; p<0,01 (рис. 7). Как и в предыдущей серии, распределение попыток проникновения в три отверстия во всех пробах серии № 2 у испытуемых экспериментальной группы отличается от аналогичного распределения у испытуемых контрольной группы (Ȥ2=12,3; df=2; p<0,01). В частности, доля неуспешных попыток проникновения в отверстие № 1 (через это отверстие испытуемые покидали пусковую камеру в предыдущей серии) за все 20 проб серии № 2 у испытуемых экспериментальной выборки составляет 44%, а у испытуемых контрольной выборки – 34%.
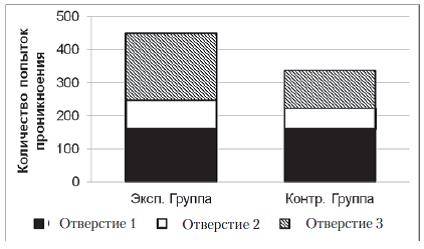
Рис. 7. Распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 3. Распределения достоверно отличаются друг от друга (Ȥ2=12,3; df=2; p<0,01)
У испытуемых экспериментальной выборки не было выявлено значимых различий в распределениях количества проникновений, совершенных в отверстия типов S, M и L, между экспериментальными сериями № 2 и № 3 (Ȥ2=1,6; df=2; p>0,05). В частности, в третьей серии не снизилась доля неуспешных попыток проникновения в отверстия типа M: в серии № 2 их доля составила 43%, в серии № 3 – 45%.
Длительные и краткие попытки проникновения в отверстия типа M наблюдались у испытуемых экспериментальной выборки в сериях № 2 и № 3. Всего за последние 2 серии у испытуемых экспериментальной выборки было зафиксировано 398 неудачных попыток проникновения в отверстия типа M. Результаты кластерного анализа приведены в табл. 2 и 3.
Таблица 2
Группа «Длительных попыток проникновения», выделенная в результате кластерного
анализа методом k-средних (в таблице приведены данные после проведения
стандартизации)
|
Переменные |
Среднее |
Станд. откл. |
DX |
|
Длительность попытки проникновения |
0,79 |
0,60 |
0,37 |
|
Глубина проникновения |
0,89 |
<0,01 |
<0,01 |
Таблица 3
Группа «Кратких попыток проникновения», выделенная в результате кластерного
анализа методом k-средних (в таблице приведены данные после проведения
стандартизации)
|
Переменные |
Среднее |
Станд. откл. |
DX |
|
Длительность попытки проникновения |
-1,00 |
0,07 |
<0,01 |
|
Глубина проникновения |
-1,12 |
<0,01 |
<0,01 |
К группе длительных попыток проникновений было отнесено 222 попытки, средняя длительность попытки проникновения – 25 с; (SD=7,61), все проникновения были глубокими – в отверстие проникала как голова змеи, так и значительная часть тела (рис. 8). К группе кратких проникновений было отнесено 176 попыток, средняя длительность попытки проникновения – 2,4 с (SD=0,90), все попытки проникновения были неглубокими – в отверстие проникала лишь часть головы змеи.
Длительные попытки проникновения преобладали в начале пробы, в то время как краткие чаще наблюдались в конце пробы (Ȥ2=64,2; df=9; p<0,01). При этом, как правило, краткие попытки совершались серией (из 3–4 попыток), идущей вслед за несколькими длительными попытками проникновения. Часто после нескольких длительных попыток проникновения змея в течение нескольких секунд быстро перемещалась по всей пусковой камере и лишь затем совершала краткие попытки проникновения. Отметим, что также краткие (по времени в несколько секунд) попытки проникновения змеи совершали и в отверстия типа S. Аналогичные попытки проникновения наблюдались и у испытуемых контрольной группы в сериях № 2 и № 3. Особенно отчетливо данная закономерность проявлялась на первых 5 пробах данных серий.
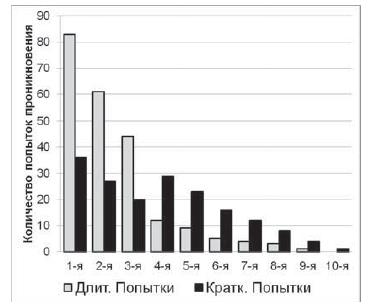
Рис. 8. Распределения частот встречаемости длительных и кратких попыток проникновения в отверстия типа M в рамках одной пробы у змей экспериментальной группы в сериях № 2 и № 3. Распределения достоверно отличаются друг от друга
Соотношения длительных и кратких попыток проникновения, совершенных в сериях № 2 и № 3 не имеют достоверных отличий друг от друга (Ȥ2=0,001; df=1; p>0,05).
Обсуждение результатов
Полученные результаты свидетельствуют, что у змей обеих групп успешно складывался навык решения экспериментальной задачи. Причем в каждой серии навык формировался заново, как при изменении границ тела, так и при изменении схемы расположения отверстий, о чем свидетельствует факт того, что в начале каждой серии у животных возрастало как время решения экспериментальной задачи, так и количество неуспешных попыток проникновения в непроницаемые отверстия.
В серии № 3 при ориентации в экспериментальной установке змеи экспериментальной выборки не научились учитывать факт изменения границ собственного тела. Навык решения экспериментальной задачи формировался у них «с нуля»: пресмыкающиеся совершали такое же количество неуспешных попыток проникновения в отверстия типа M, как и в предыдущей серии.
Однако прежде чем переходить к выводам, обсудим соотношение длительных и кратких попыток проникновения в отверстия типа M, продемонстрированных испытуемыми экспериментальной группы в сериях № 2 и № 3. Тот факт, что первые (в рамках каждой пробы) попытки проникновения чаще являлись длительными, а завершающие пробу попытки – напротив, краткими, является свидетельством динамики научения змей в рамках каждой отдельной пробы. Иначе говоря, к концу пробы змеи тратили меньше времени и усилий на попытки проникновения в непроницаемые для них отверстия типа M. Мы полагаем, что этот факт свидетельствует о том, что к концу каждой пробы у змей происходила определенная перестройка психического отражения собственного события со средой проблемной ситуации.
Необходимо обратить внимание, что сходные длительные и краткие попытки проникновения наблюдались в эксперименте по изучению способности крыс учитывать изменение границ собственного тела (Хватов и др., 2014b). В этом эксперименте крысам также необходимо было проникать мордой в различные по диаметру отверстия (трех типов, аналогичных тем, что использовались в настоящем эксперименте, с учетом размеров тела крысы) для достижения приманки. Эксперимент состоял из 5 серий: увеличение границ тела животных было произведено перед началом 2-й серии, а в следующих трех сериях происходило варьирование расположения отверстий в экспериментальной установке. У крыс длительные попытки проникновения, сопровождавшиеся «буксующими» движениями лап в направлении отверстия, за весь эксперимент наблюдались только на первых 5 пробах серии № 2 (всего в серии было также 20 проб), количество кратких попыток проникновения оставалось стабильным на протяжении всех последующих проб. Этот факт был объяснен тем, что в отличие от длительных попыток проникновения, при осуществлении кратких попыток, крыса не стремилась проникнуть в отверстие и достать приманку, но лишь кинестетически обследовала отверстие, определяя его диаметр, а также обнаруживала место расположения приманки. Соответственно, краткие проникновения осуществляли только ориентировочную функцию. Мы полагаем, что в нашем эксперименте у змей краткие попытки проникновения выполняли эту же функцию. Однако если у крыс происходило изменение схемы собственного тела, что выражалось в том, что грызуны более не совершали длительных попыток проникновения в отверстия, непроницаемые для их увеличенного тела, даже при изменении схемы экспериментальной установки, то у змей, как мы уже отмечали, в серии № 3 подобного переноса опыта обнаружено не было. Напротив, у пресмыкающихся соотношения длительных и кратких попыток проникновения в сериях № 2 и № 3 не имеют существенных отличий друг от друга.
В более ранних исследованиях способности учитывать параметры собственного тела членистоногими – тараканами Рeriplaneta americana (Хватов, 2011) и Nauphoeta cinerea, а также сверчками Gryllus assimilis (Хватов и др., 2013b), в которых в экспериментальной группе животных также увеличивались границы тела насекомых, а в контрольной изменялась схема отверстий в экспериментальной установке, были получены совершенно иные результаты. Во-первых, неуспешные попытки проникновения в ранее проницаемые отверстия у этих насекомых не удалось разделить на длительные и краткие. Во-вторых, членистоногие, как при изменении границ тела, так и при изменении размера отверстий, демонстрировали одинаковое поведение. У змей же увеличение границ тела вызвало большие затруднения, нежели изменение диаметра отверстий, ранее являвшихся проницаемыми. Об этом свидетельствуют большие временные промежутки, затраченные на решение задачи, и большее количество неудачных попыток проникновения в отверстия № 1 и № 3 в сериях № 2 и № 3 соответственно, наблюдавшиеся у испытуемых экспериментальной группы в сравнении с испытуемыми контрольной группы.
Обозначим различия в поведении вышеописанных групп животных при решении задачи на учет физических параметров собственного тела (табл. 4).
Таблица 4
Сопоставление способов решения задачи на способность учитывать изменение границ
собственного тела у различных животных
|
|
Животные |
||
|
Критерии сопоставления |
Насекомые |
Крысы |
Змеи |
|
Обнаруживаются ли различия при решении задачи в случае изменения границ тела и в случае изменения характеристик внешней среды? |
Нет |
Да |
Да |
|
Выделяются ли краткие и длительные попытки проникновения? |
Нет |
Да |
Да |
|
Отсутствуют ли длительные попытки проникновения при последующем изменении характеристик внешней среды? |
– |
Да |
Нет |
С позиции разрабатываемой нами концепции самоотражения животных и человека (Хватов, 2010; 2014) возникшую в результате подобного сравнения картину можно проинтерпретировать следующим образом. В психике насекомых схема тела в качестве самостоятельной автономной структурной единицы еще не выделяется. Однако осуществлять какую бы то ни было поведенческую активность без учета характеристик собственного тела невозможно. У насекомых отражение характеристик собственного тела существует в неразрывном единстве с отражением внешних условий (схемы расположения отверстий в экспериментальной установке). Можно утверждать, что субъективно они воспринимают лишь собственную «неспособность» проникнуть в отверстие, но не воспринимают, чем обусловлена эта неспособность: изменениями их собственного тела или изменениями внешней среды. Поэтому, сталкиваясь с такой «неспособностью», насекомые полностью перестраивают психическое отражение собственного события с внешним миром вместе с восприятием собственного тела и восприятием внешней среды, т.е. выстраивают полностью новый навык – «с нуля».
С другой стороны, у крыс схема тела уже обособляется в самостоятельный когнитивный конструкт и, будучи сформирована в рамках одного события (в рамках связи с одними характеристиками внешней среды), может быть экстраполирована за его пределы и включена в рамки других событий (в связи с другими характеристиками внешней среды). Соответственно, у крыс в сравнении с членистоногими психический образ имеет большую дифференцированность и гибкость организации, что позволяет использовать элементы ранее сформированных навыков для выстраивания новых и благодаря этому экономить время и совершать меньше ошибок. Это соответствует общему представлению о направлениях эволюции психики у представителей разных эволюционных таксонов (Хватов, 2012a; 2012b).
Психика змей, при данном ракурсе рассмотрения, по своей форме организации представляет собой промежуточный вариант между психикой членистоногих и психикой крыс (млекопитающих). Во-первых, змеи не только отражают собственную «неспособность» проникнуть в отверстие, но также по-разному реагируют на то, из-за чего возникает эта «неспособность»: за счет увеличения границ тела или за счет изменений диаметра отверстий в экспериментальной установке. Во-вторых, схема тела этих пресмыкающихся, хотя и довольно тесно слита с отражением конкретных внешних условий, все же может быть экстренно модифицирована, что, соответственно, ведет к изменению поведения. Однако такая модификация осуществляется лишь в рамках весьма узкой (по времени и пространству) событийной ситуации – в нашем случае речь идет об отдельной экспериментальной пробе. Перенос этих изменений схемы тела в более отдаленные событийные ситуации (в другие экспериментальные пробы) оказывается невозможным.
Полученные в настоящем исследовании результаты показывают, что у змей, как представителей класса пресмыкающихся, обнаруживаются предпосылки дифференциации схемы собственного тела и схемы внешней среды в рамках целостного психического отражения организмом собственного события с внешней средой. Этот процесс дифференциации является магистральной линией развития самоотражения у позвоночных и в наиболее явной форме обнаруживается у млекопитающих и птиц. Полученные результаты нуждаются в дополнительной проверке на материале поведения других пресмыкающихся в аналогичных экспериментальных условиях.
Выводы
Мы можем заключить, что гипотеза исследования была подтверждена частично. С одной стороны, змеи оказались способны модифицировать собственную схему тела, что выражалось в изменении их поведения с учетом увеличенных границ собственного тела путем научения. С другой стороны, поскольку у змей схема собственного тела неотделима от их восприятия предметного контекста ситуации, формирование нового навыка обязательно требует перестройки и восприятия внешней среды и восприятия собственного тела.
Финансирование
Работа поддержана грантом РГНФ № 14-06-00669, тип проекта – а.Литература
- Барабанщиков В. А. Восприятие и событие. СПб.: Алетейя, 2002. 512 с.
- Столин В.В. Самосознание личности. М.: Издательство Московского Университета, 1983. 284 с. Хватов И. А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 1 [Электронный ресурс] // Психологические исследования. 2012а. № 1 (21). С. 1. URL: http://psystudy.ru/index.php/num/2012n1-21/622-khvatov21.html (дата обращения: 22.09.2014). doi:0421200116/0001.
- Хватов И. А. Главные направления эволюции психики в контексте онтологического и дифференционно-интеграционного подходов. Часть 2 [Электронный ресурс] // Психологические исследования. 2012b. № 2 (22). С. 12. URL: http://psystudy.ru/index.php/num/2012n2-22/648-khvatov22.html (дата обращения: 22.09.2014). doi:0421200116/0024.
- Хватов И. А. Особенности самоотражения у животных на разных стадиях филогенеза: дисс. ... канд. психол. наук. М., 2010.
- Хватов И. А. Проблема интеллекта животных в контексте структурно-интегративного и дифференционно-интеграционного подходов [Электронный ресурс] // Психологические исследования. 2013. Т 6. № 28. С. 1. URL: http://psystudy.ru/index.php/num/2013v6n28/788-khvatov28.html (дата обращения: 28.07.2014).
- Хватов И. А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011. Т 4. № 1. C. 28-39.
- Хватов И. А. Эволюция самоотражения животных и человека в контексте дифференционно-интеграционного подхода // Дифференционно-интеграционная теория развития. Кн. 2. / Сост. и ред. Н.И. Чуприкова, Е.В. Волкова. М.: Языки славянских культур, 2014. С. 343-360.
- Хватов И. А., Соколов А.Ю, Харитонов А.Н. Методика изучения схемы тела у змей // Экспериментальная психология. 2015. Т. 8. № 1. С. 85-93.
- Хватов И.А., Соколов А.Ю, Харитонов А.Н., Куличенкова К.Н. Восприятие собственного тела у крыс // Естественно-научный подход в современной психологии / Отв. ред. В. А. Барабанщиков. М.: Институт психологии РАН, 2014а. С. 326-332.
- Хватов И. А., Соколов А. Ю., Харитонов А. Н., Куличенкова К. Н. Методика изучения схемы тела у мелких млекопитающих // Экспериментальная психология. 2014b. Т 7. № 3. С. 137-144.
- Хватов И. А., Харитонов А. Н. Модификация плана развертки собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica // Экспериментальная психология. 2013. Т 6. № 2. С. 101-114.
- Хватов И. А., Харитонов А. Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. Т 5. № 3. C. 96-107.
- Хватов И. А., Харитонов А. Н., Соколов А. Ю. Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Gryllus assimilis // Экспериментальная психология. 2013a. Т. 6. № 4. С. 79-95.
- Хватов И.А., Харитонов А.Н., Соколов А.Ю. Специфика восприятия собственного тела при регуляции поведения у беспозвоночных и позвоночных: эволюционный и сравнительный аспекты // Шестая международная конференция по когнитивной науке: Тезисы докладов / Под общ. ред. Б. Величковского, В. Рубцова, Д. Ушакова. Калининград, 2014. С. 603-604.
- Хватов И.А., Харитонов А.Н, Соколов А.Ю. Самоотражение у беспозвоночных // Эволюционная и сравнительная психология в России: традиции и перспективы / Под ред. А. Н. Харитонова. М.: Институт психологии РАН, 2013b. С. 170-177.
- Asendorpf J.B., Warkentin V, Baudonniere P.M. Self-Awareness and Other-Awareness II: Mirror SelfRecognition, Social Contingency Awareness, and Synchronic Imitation // Developmental Psychology. 1996. Vol. 32. № 2. P 313-321.
- Bekoff M. Observations of scent-marking and discriminating self from others by a domestic dog (Canis familiaris): tales of displaced yellow snow // Behavioural Processes. 2001. Vol. 55. № 2. P 75-79.
- Gallagher S., Cole J. Body Schema and Body Image in a Deafferented Subject // Journal of Mind and Behavior. 1995. Vol. 16. P 369-390.
- Gallup G.Jr. Chimpanzees: Self recognition // Science. 1970. Vol. 167. № 3914. P 86-87.
- Herman L.M., Matus D. S., Herman E. Y.K., Ivancic M., Pack A. A. The bottlenosed dolphin’s (Tursiops truncatus) understanding of gestures as symbolic representations of its body parts // Animal Learning & Behavior. 2001. Vol. 29. № 3. P 250-264.
- Maravita A., Iriki A. Tools for the body (schema) // Trends in Cognitive Sciences. 2004. Vol. 8. № 2. P 79-86.
Информация об авторах
Метрики
Просмотров
Всего: 4520
В прошлом месяце: 34
В текущем месяце: 2
Скачиваний
Всего: 921
В прошлом месяце: 4
В текущем месяце: 0