ЛЕВЕНЕЦ Я. В. , Институт систематики и экологии животных СО РАН, Новосибирск, Россия, e-mail: jan.levenets@gmail.com
ПАНТЕЛЕЕВА С. Н.[*] [†], Институт систематики и экологии животных СО РАН; Новосибирский государственный университет, Новосибирск, Россия, e-mail: psofia@mail.ru
РЕЗНИКОВА Ж. И.[‡], Институт систематики и экологии животных СО РАН; Новосибирский государственный университет, Новосибирск, Россия, e-mail: zhanna@reznikova.net
На примере охотничьего поведения мелких млекопитающих проведен сравнительный межвидовой анализ стереотипов, включающих элементы гибкого поведения. Исследовались реакции на подвижную добычу (тараканов) у грызунов с различной пищевой специализацией в сравнении с представителем отряда насекомоядных (обыкновенной бурозубкой). Статистический анализ данных проводился на основании количественного анализа этограмм, а для визуализации схем поведенческих стереотипов рассчитывались матрицы вероятностей перехода между элементами поведения (цепи Маркова первого порядка). Отличительной особенностью стереотипа грызунов является следующий за захватом зубами захват и удержание добычи передними лапами. Данный способ захвата является более эволюционно прогрессивным, чем захват добычи только зубами, как происходит у бурозубок. Схемы стереотипов и тактики охоты зеленоядной узкочерепной полевки и зерноядной полевой мыши оказались схожи со схемой охотничьего поведения у специализированного представителя насекомоядных - обыкновенной бурозубки. Стереотип охотничьего поведения крысы наименее специализирован. У исследованных видов грызунов стереотип более примитивный, чем у специализированных хищных грызунов, но обладает чертами высокой специфичности и может рассматриваться как поведенческая адаптация, позволяющая расширить спектр пищевых ресурсов путем активной охоты на насекомых. Предложенная схема сравнительного анализа стереотипного поведения может использоваться не только в сравнительно-этологических, но и в психологических исследованиях.
Введение
История изучения поведенческих стереотипов уходит корнями в психиатрию, сравнительную психологию и раннюю генетику поведения. Одно из ранних и наиболее известных исследований в этой области принадлежит британскому психиатру Джону Лэнгдону Дауну (Down, 1887), который классифицировал поведенческие паттерны (стереотипы), характерные для идиотов разных этнических групп. «Монголоидные» стереотипы позже были описаны как «синдром Дауна». В начале XX в. психиатры обратили внимание на то, что при некоторых психических заболеваниях люди избирают тактику навязчиво повторяющихся действий, напоминающих ритуализованное поведение животных (Berrios, 2011). Так, одна из характеристик, общая для расстройств аутического спектра, состоит в крайней ритуализации действий. Психиатры характеризуют такое поведение как стремление к повторению и предохраняющее поведение (Bishop, Richler, Lord, 2006).
Однако стереотипизация поведения, которая для человека в раннем возрасте может находиться на грани нормы, а в более зрелом возрасте сигнализировать о патологии, для огромного числа видов животных является частью видового репертуара. Впервые понятие «канализованного поведения» было предложено Эдвином Хольтом (Holt, 1933), который обратил внимание на развитие специфических сенсомоторных путей, «возникающих из хаоса» на ранних стадиях эмбрионального развития (обзор см.: Gottlieb, 1991). На основе своих первых данных по генетике поведения Конрад Уоддингтон (Waddington, 1942, 1968) предложил образную концепцию «эпигенетического ландшафта», согласно которой организм проходит в своем развитии предопределенную сочетанием генетических и средовых факторов траекторию («креод»). Уоддингтон был первым, кто сформулировал современное представление о роли мутаций как факторов, влияющих на процесс развития организма и, следовательно, составляющих основу изменчивости - одного из трех главных факторов биологической эволюции. Что касается изучения поведенческих стереотипов, то исследование изменчивости приобретает особую важность с точки зрения формирования представлений об эволюционных основах поведения (Зорина, Полетаева, Резникова, 2013; Резникова, 2016).
К настоящему времени установлен факт отсутствия сколь-нибудь надежных критериев, позволяющих разделить составляющие инстинкта и обучения в поведении животного, разработка и применение которых возможна вне рамок проведения специальных экспериментов (Bateson, Gluckman, 2011). Выученные цепочки действий у животных могут стать настолько рутинными и жестко повторяющимися, что по своим проявлениям напоминают цепочки инстинктивных действий (Резникова, Пантелеева, 2015). В этологии и сравнительной психологии под поведенческими стереотипами понимают поведенческие последовательности, состоящие из устойчиво повторяющихся элементов (Пантелеева, Данзанов, Резникова, 2010). От стереотипов отличаются стереотипии: повторяющиеся неизменные формы поведения без очевидной цели и функции (Mason, 1991). Не только у людей, но и у животных стереотипии указывают на патологию в поведении. Они не встречаются у животных в естественной среде и характерны либо для специально выведенных генетических линий (Francis et al., 2003), либо для животных в неволе, причем они настолько явно свидетельствуют о существенных недостатках в их содержании, что используются как диагностический признак (Broom, Johnson, 1993). Стереотипы, в отличие от стереотипий, не повторяются циклически; поведенческая цепочка имеет начало и завершение, однако границы между этими формами поведения могут быть и размытыми (Резникова, Пантелеева, Левенец, 2014; Reznikova, Panteleeva, Danzanov, 2012).
Сравнительное исследование стереотипов, одинаково направленных у разных видов животных, позволяет развить представление об эволюционном формировании и путях адаптации поведения (Зорина, Полетаева, Резникова, 2013; Резникова, 2016).
Хорошим модельным примером служит стереотип охотничьего поведения у грызунов. Среди них представлены как специализированные охотники - представители рода кузнечиковых хомячков Onychomys, обладающие морфологическими и физиологическими адаптациями (Sarko et al., 2011), так и эврифаги, проявляющие охотничье поведение факультативно, например, оленьи хомячки (Peromyscus maniculatus) (Langley, 1994). Недавно обнаруженное охотничье поведение у полевых мышей (Apodemus agrarius) (Panteleeva, Reznikova, Vygonyailova, 2013) и рыжих полевок (Myodes glareolus) (Konczal et al., 2016), не имеющих морфологических или физиологических адаптаций к охоте, позволяет предположить наличие у них специфических поведенческих приспособлений. Возникает вопрос о наличии у грызунов стереотипов поведения, использование которых расширяет адаптивный потенциал вида за счет возможности переключения на животную пищу.
Цель данной работы - на примере охотничьего поведения мелких млекопитающих провести сравнительный межвидовой анализ стереотипов, включающих элементы гибкого поведения. Для этого мы исследовали реакции на подвижных насекомых у грызунов с различной пищевой специализацией в сравнении с представителем насекомоядных (обыкновенной бурозубкой).
Материалы и методы
Исследования проводились в 2012-2015 гг. в лаборатории на серой крысе (Rattus norvegicus) (n=81), известной как всеядный грызун, зерноядной полевой мыши (A. agrarius) (n=26), зеленоядной узкочерепной полевке (Lasiopodomys gregalis) (n=43) и обыкновенной бурозубке (Sorex araneus) (n=11). Все особи серой крысы и 9 полевых мышей родились в лаборатории и до тестирования не имели опыта охоты, остальные животные были отловлены в естественных условиях (Новосибирская область). Зверьки содержались в индивидуальных клетках и имели постоянный доступ к воде и пище.
Для наблюдений животных по одному помещали в прозрачную арену Noldus (45^45x50 см для крыс, 30x30x35 см для остальных). Спустя 5 мин в арену помещали добычу - мраморного таракана (Nauphoeta cinerea) (средняя длина тела 27,93±0,22 мм). Если животное охотилось, то на арену помещали последовательно еще двух тараканов, если не проявляло интереса к добыче, то наблюдения прекращали спустя 10 минут. Серые крысы тестировались один раз. Поскольку особей остальных видов было меньше, чем крыс, то для получения сопоставимого количества стереотипов было увеличено количество тестов: полевых мышей и обыкновенных бурозубок тестировали два раза, а узкочерепных полевок - три. Действия животных фиксировались с помощью видеокамер: Sony Handycam DCR- SR68 (крысы и мыши), Sony HDR-AS200V (полевки и бурозубки). Анализировались только случаи успешной охоты, закончившиеся поимкой и поеданием добычи.
Для описания поведения в качестве элементарной единицы нами выделяются элементарные двигательные акты и позы («элементы поведения»). Поведенческой последовательностью мы называем произвольный набор последовательно совершаемых элементов поведения. Внутри поведенческих последовательностей мы выделяем поведенческие стереотипы, состоящие из устойчиво повторяющихся «цепочек» элементов поведения (подробно см.: Резникова, Пантелеева, Левенец, 2014). Всего было выделено 19 элементов поведения, разделенных на 3 типа. «Ключевые», без которых совершение стереотипа невозможно: преследование добычи бегом (Q) или спокойным шагом (S), укус (W), захват добычи лапами (E) (только у грызунов). «Дополнительные» элементы («приготовления» к охоте и поеданию добычи) присутствовали не во всех стереотипах: принюхивание (D), перенос добычи в зубах (G), перехват (R) (только у грызунов), откусывание конечностей добычи (H) и придерживание добычи одной (N) или двумя лапами (M) - только у бурозубки. «Шумовые элементы» (не влияющие на совершение стереотипа): замирание (C), поворот корпуса на 90° (V), разворот корпуса 180° (B), поворот головы (F), вертикальная стойка (I), стойка с опорой на арену (Y), движения назад (U), прыжок (J) и чистка (X). Некоторые ключевые элементы поведения представлены на рис. 1.

Рис. 1: а) обыкновенная бурозубка кусает добычу (W - «укус»); б) узкочерепная полевка откусывает конечности добычи (H - «откусывание конечностей»); В) полевая мышь захватывает лапами пойманную зубами добычу (W-E «укус - захват лапами»); Г) серая крыса удерживает пойманную добычу в передних лапах
Обработка видеозаписи проводилась с 25-кратным замедлением в программе The Observer XT 10 (Noldus Information Technology). Используя полученный «алфавит» из 16 элементов, мы преобразовали демонстрируемое поведение в последовательности букв, где каждая буква соответствовала одному элементу поведения. Полученные поведенческие последовательности охотничьих стереотипов перемещались в отдельные для каждого вида «суммарные» текстовые файлы (в формате txt).
Сравнения долей охотившихся и не охотившихся особей у разных видов, а также успешных и неуспешных атак у разных особей проводились с помощью точного теста Фишера. Анализ количества элементов в стереотипах проведен с помощью H-критерия Краскела-Уоллиса; при сравнении представлены медиана, первый и третий квартили (Me; Q1 - Q3). Для построения схемы стереотипа, используя «суммарные» текстовые файлы, мы рассчитывали матрицы вероятностей перехода от одного поведенческого элемента к другому (Марковский процесс первого порядка) (Casarrubea, Sorbera, Crescimanno, 2008).
Результаты
При первом предъявлении подвижной добычи охотничье поведение продемонстрировали 67,9% (55 из 81) особей серой крысы, 65,4% (17 из 26) полевых мышей, 18,5% (9 из 46) узкочерепных полевок и 100% обыкновенных бурозубок (n=11). Полевки проявляли охотничье поведение достоверно реже, чем крысы, мыши и бурозубки (p < 0,001 для всех случаев). Всего было зафиксировано 125 успешных и 77 неуспешных случаев охоты у крыс, 83 и 39 - у полевых мышей, 34 и 92 - у полевок, 61 и 34 - у бурозубок соответственно. Успешность атак у узкочерепной полевки оказалась ниже, чем у всех остальных (p < 0,001 для всех случаев).
По длине охотничьи стереотипы у серых крыс (16; 9 - 28), полевок (9; 5 - 43) и бурозубок (22; 13 - 34) достоверно не различались между собой (H = 5,5, NS), но были короче, чем у мышей (31; 19 - 56) (H = 35,1; p < 0,01).
Мы сравнили скорость охоты у разных видов (отношение между длиной стереотипа и его продолжительностью). Обыкновенные бурозубки демонстрировали 2,9 (1,8 - 4,2) элементов поведения в секунду, что достоверно выше, чем у серых крыс (1,2; 0,9 - 1,5), полевых мышей (2,1; 1,5 - 2,8) и полевок (1,6; 1,2 - 2,2) (H = 103,9; p < 0,01). Скорость охоты у полевых мышей была выше, чем у узкочерепных полевок (H = 7,3; p < 0,01), а у серых крыс этот показатель оказался наименьшим (H = 103,9; p < 0,01).
Схемы стереотипов охотничьего поведения представлены на рис. 2, где показаны все устойчивые связи между элементами (вероятность перехода от одного элемента к другому p > 0,2) и некоторые неустойчивые (р < 0,2), но важные для совершения стереотипа связи.
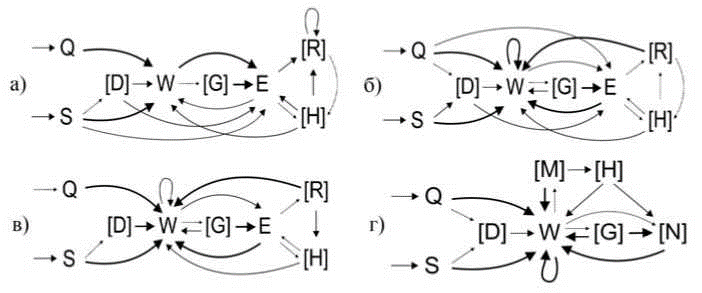
Рис. 2. Схемы охотничьих стереотипов серой крысы (а), полевой мыши (б), узкочерепной полевки (в) и обыкновенной бурозубки (г). Тонкой пунктирной линией обозначены некоторые неустойчивые связи между элементами (р < 0,2). Простой линией обозначены устойчивые связи (0,2 < р < 0,5). Жирной линией обозначены высоко устойчивые связи между элементами (p > 0,5). В квадратных скобках указаны дополнительные элементы
У всех исследованных видов охотничий стереотип, как правило, начинался с преследования добычи бегом (Q) или спокойным шагом (S), а в тех редких случаях, когда таракан сам приближался слишком близко, фаза преследования могла отсутствовать. Затем могло следовать принюхивание (D), и совершался захват зубами («укус», один или несколько подряд, W). С этого момента схемы стереотипов грызунов и обыкновенной бурозубки расходятся. Грызуны после «укуса» захватывают добычу обеими передними конечностями (Е) для поедания. У бурозубок этот элемент поведения отсутствует, они либо сразу схватывают добычу зубами, либо одной (N) или двумя (M) передними лапами прижимают насекомое ко дну арены, продолжая совершать серию быстрых укусов. В охотничьих стереотипах серых крыс после захвата добычи лапами чаще встречались перехваты (R) - манипуляции с удерживаемой в лапах добычей (повороты, перевороты). Данное поведение очень редко наблюдалось у мышей и полевок и никогда не отмечалось у бурозубок. Схватив добычу, зверьки могли либо сначала откусывать конечности насекомого (H), либо сразу перейти к его поеданию. Элемент поведения, предшествующий поеданию, выступает в качестве окончания успешного охотничьего стереотипа. У грызунов такими финальными элементами могли быть: укус (W), захват лапами (E), перехват (R) или откусывание конечностей добычи (H); а у бурозубок: укус (W), придерживание добычи одной лапой (N), откусывание конечностей добычи (H), а также принюхивание (D) и перенос добычи в зубах (G), встречавшиеся единично.
Результаты количественного анализа представлены на рис. 3. Стереотипы охотничьего поведения полевой мыши содержат наибольшее количество ключевых элементов поведения «укус» (20; 10-30) (H = 102,5, p < 0,01). У полевок (5,5; 5-21) и у крыс (5; 5-8) этот показатель достоверно не различается (H = 0,4, NS), но был меньше, чем у мышей и бурозубок (15; 8 - 22) (H = 102,5, p < 0,01). На один стереотип полевок приходится меньше элементов поведения «захват лапами» (2; 1-8,5), чем у крыс (5; 3-10) и мышей (7; 4-11) (H = 66,3, p < 0,01), при этом у мышей этот показатель выше, чем у крыс (H = 4,9, p < 0,05). Наибольшее количество дополнительных элементов поведения «перехват добычи» наблюдается в стереотипах крыс (3; 1-4) (H = 78,5, p < 0,01), а наименьшее - у полевок. Количество актов откусывания конечностей пойманной добычи в стереотипах мышей (1; 0-2), полевок (1; 0-2,8) и бурозубок (1; 0-3) достоверно не различается (H = 0,8, NS), а у крыс этот показатель - наименьший (0; 0-1) (H = 26,3, p < 0,01).
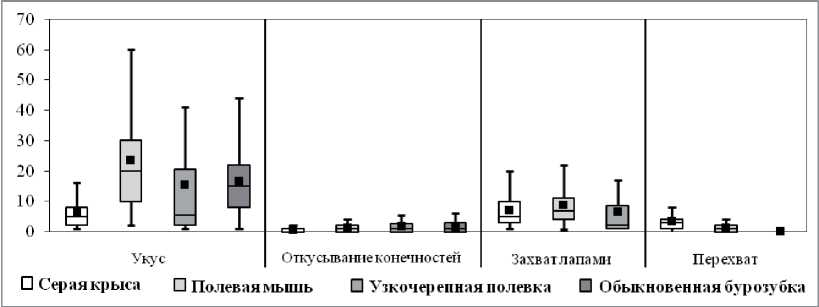
Рис. 3. Количество элементов поведения, приходящихся на один успешный стереотип охотничьего поведения у разных видов
Далее в зависимости от их характера все укусы были разделены на 3 группы: одиночные, двойные и множественные (3 и более укусов подряд). Отличительная особенность стереотипов крыс в том, что в них содержались только одиночные укусы, в то время как в стереотипах мышей, полевок и бурозубок присутствовали двойные и множественные укусы (рис. 4). В охотничьих стереотипах бурозубок количество одиночных укусов наименьшее (1; 0-3) (H = 26,3; p < 0,01) и достоверно отличается от практически одинаковых значений этого показателя в стереотипах крыс (4; 2-7), мышей (4; 3-7) и полевок (2; 1-7) (H = 5,8, NS). Количество двойных укусов в стереотипах мышей (1; 0-3), полевок (1; 0-3) и бурозубок (1; 0-1) достоверно не различалось (H = 5,1, NS). Количество множественных укусов у полевок (0; 0-0,8) было достоверно меньше, чем у мышей (4; 1-4) и у бурозубок (2; 1-3) (H = 19,2; p < 0,01).
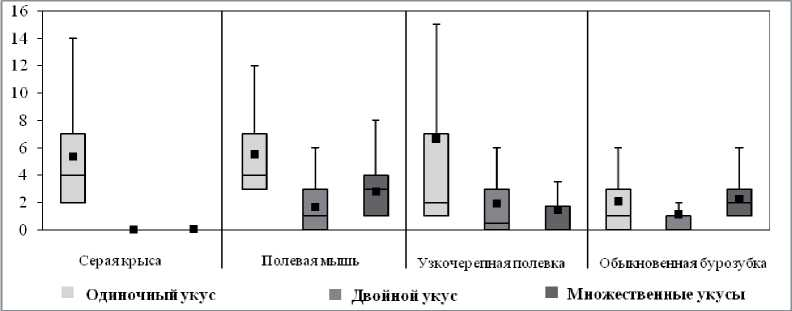
Рис. 4. Количество одиночных, двойных и множественных укусов, приходящихся на один успешный стереотип охотничьего поведения у разных видов животных
Обсуждение и заключение
Количественное сравнение одинаково направленных стереотипов у разных видов животных позволило выявить охотничье поведение, ранее неизвестное для исследуемых видов, и сделать предположение о характере поведенческих адаптаций к плотоядному образу жизни у грызунов. Внимание исследователей к факультативному охотничьему поведению зерноядных грызунов было привлечено совсем недавно, а зеленоядные в этом плане практически не исследовались, за исключением нескольких видов эврифагов, в различной степени тяготеющих к питанию растительной пищей (Timberlake, Washburn, 1989). Эффективное охотничье поведение у зерноядных грызунов с детальным описанием стереотипа охоты было впервые выявлено у полевой мыши (Panteleeva, Reznikova, Vygonyailova, 2013). На примере другого, столь же широко распространенного и многочисленного зерноядно- зеленоядного вида - рыжей полевки (M. glareolus) - было показано, что отбор успешных «охотников на насекомых» в лаборатории приводит к быстрому, в течение нескольких поколений, повышению частоты проявления охотничьего поведения, что служит примером «быстрой эволюции» (Konczal et al., 2016). Характер охотничьего поведения в этой работе описан не был. Мы впервые провели детальное сравнение охотничьего поведения у представителей зерноядных (полевая мышь), зеленоядных (узкочерепная полевка) и всеядных (серая крыса) грызунов с насекомоядным видом, облигатно проявляющим охотничье поведение, - обыкновенной бурозубкой.
У зеленоядной узкочерепной полевки оказалась самая меньшая среди исследованных видов доля охотящихся особей и наиболее низкая успешность охоты. Примечательно, однако, что у тех особей, которые охотились, схема охотничьего стереотипа практически не отличалась от схемы стереотипа у полевых мышей. Это означает, что в поведенческом репертуаре зеленоядных полевок есть полный и специфический стереотип охотничьего поведения, типичный для охотящихся грызунов.
У исследованных грызунов схемы охотничьего стереотипа по порядку совершения ключевых элементов оказались сходными, как и сам характер охотничьего поведения. По- видимому, это поведение не зависит от размеров добычи. В наших предварительных экспериментах было выяснено, что полевые мыши и узкочерепные полевки атакуют мелкую малоподвижную добычу (личинок мучного хрущака) точно так же, как это делают серые крысы при охоте, например, на мраморного таракана: сближение с добычей, укус, затем захват в передние лапы (Резникова, Пантелеева, Левенец, 2014).
Существенное различие между охотничьими атаками грызунов и насекомоядных состоит в том, что грызуны после захвата добычи зубами («укуса») осуществляют захват лапами, а бурозубки используют только зубы. Примечательно, что бурозубки атакуют добычу любых размеров только быстрыми укусами, что оценивается как более примитивный способ по сравнению с захватом лапами - относительно недавним достижением в филогенетическом плане (Eisenberg, Leyhausen, 1972). Одним из показателей большей специализации кузнечикового хомячка, как грызуна-хищника, является начало атаки с захвата добычи лапами (Langley, 1994). В нашем исследовании полевые мыши и узкочерепные полевки, в отличие от кузнечиковых хомячков, начинали атаку с захвата насекомого зубами (укуса), а затем хватали его лапами.
Результаты сравнительного анализа показателей длины стереотипов и скорости охоты характеризуют бурозубку как эффективного специализированного хищника. Серая крыса также естественно выступает как хищник-генералист, не проявляющий специализированного охотничьего поведения. Крысы используют укус для удержания добычи и последующего захвата лапами, они активно манипулируют удерживаемой добычей, но не умерщвляют ее, а поедают живьем, поэтому добыча часто вырывается и зверек вынужден возобновлять преследование. Примечательно, что как полевые мыши, так и узкочерепные полевки действуют со значительно большей скоростью и гораздо эффективнее, чем крысы, повреждая и умерщвляя добычу серией укусов, так же, как это делают бурозубки.
На основании классификации охотничьего поведения мелких грызунов (Langley, 1994) можно предположить, что не только зерноядные полевые мыши, но и зеленояд- ные узкочерепные полевки обладают прогрессивными чертами охотничьего стереотипа по сравнению с насекомоядными, т. е. имеют поведенческие адаптации к плотоядному образу жизни. Охотничий стереотип у этих видов более примитивный, чем у специализированных хищных грызунов, но обладает чертами высокой специфичности и может позволить им расширить спектр пищевых ресурсов путем активной охоты на насекомых. Предложенная схема сравнительного анализа стереотипного поведения может использоваться не только в сравнительно-этологических, но и в психологических исследованиях.
[†] Пантелеева С. Н. Кандидат биологических наук, старший научный сотрудник, Институт систематики и экологии животных СО РАН (ФБГУН ИСиЭЖ СО РАН); Новосибирский государственный университет (ГБОУ ВПО НГУ). E-mail: psofia@mail.ru
[‡] Резникова Ж. И. Доктор биологических наук, профессор, ведущий научный сотрудник, Институт систематики и экологии животных СО РАН (ФБГУН ИСиЭЖ СОРАН); Новосибирский государственный университет (ГБОУ ВПО НГУ). E-mail:, zhanna@reznikova.net