Экспериментальная психология
2022. Том 15. № 3. С. 121–139
doi:10.17759/exppsy.2022150309
ISSN: 2072-7593 / 2311-7036 (online)
Успешность распознавания лицевых экспрессий носителями различных генотипов генов COMT, DRD4, 5HT2A, MAOA
Аннотация
Общая информация
Ключевые слова: лицевые экспрессии, эмоции, распознавание эмоций, распознавание эмоций по лицевым экспрессиям, мимика
Рубрика издания: Наука о лице
Тип материала: научная статья
DOI: https://doi.org/10.17759/exppsy.2022150309
Финансирование. Исследование выполнено при финансовой поддержке Российского научно- го фонда (РНФ) в рамках научного проекта №20-64-47057.
Получена: 08.06.2022
Принята в печать:
Для цитаты: Ковш Е.М., Явна Д.В., Бабенко В.В., Ермаков П.Н., Воробьева Е.В., Денисова Е.Г., Алексеева Д.С. Успешность распознавания лицевых экспрессий носителями различных генотипов генов COMT, DRD4, 5HT2A, MAOA // Экспериментальная психология. 2022. Том 15. № 3. С. 121–139. DOI: 10.17759/exppsy.2022150309
Полный текст
Введение
Успешность распознавания лицевых экспрессий связана с визуальным и социальным познанием и может изучаться в контексте разных подходов и разделов науки. На сегод- няшний день широко исследуются социокультурные и психологические корреляты успеш- ности распознавания лицевых экспрессий: с позиций гуманитарных наук рассматриваются вопросы связи данного феномена с эмоциональным и культурным интеллектом, когнитив- ными стилями, этнической принадлежностью, опытом общения и другими социально-ин- теллектуальными особенностями [1; 4; 5]. Рядом авторов показано, что распознавание лиц имеет особенное значение в социальном взаимодействии, и нарушения этой способности часто встречаются в контексте разных клинических синдромов: расстройств аутистиче- ского спектра, тревожных расстройств и др. [10; 27; 43]. На современном этапе изучение проблемы распознавания лицевых экспрессий приобрело особенное, прикладное значение ввиду возможности применения полученных результатов для разработки современных компьютерных программ и приложений [7; 9].
На сегодняшний день выявлено, что на успешность распознавания лицевых экс- прессий оказывает влияние целый ряд факторов: наследственность, морфофункциональ- ные особенности головного мозга, особенности культурной среды, семейный опыт и др. Согласно модели нейроповеденческого созревания, различия в обработке мимических про- явлений эмоций могут быть связаны с унаследованными особенностями функционирова- ния «эмоциональных» нейронных схем, к числу которых можно отнести миндалевидное тело и другие лимбические структуры [32]. В целом, в качестве мозговых механизмов рас- познавания лицевых экспрессий на сегодняшний день принято рассматривать интеграцию работы затылочной коры, веретенообразной извилины, миндалины, дорсолатеральной и вентромедиальной префронтальной коры. К примеру, обработка грустных и испуганных выражений лица сопровождается активацией связи между миндалиной и дорсолатеральной областью коры головного мозга [18]. При этом стоит отметить, что на процесс восприятия и регуляции эмоций оказывают влияние ситуационные (динамичные во времени—стресс, усталость, голод, мотивация) и диспозиционные факторы (черты личности, темперамент) [22]. Исследователями показано, что людям с депрессией требуется больше времени для распознавания положительных, но не отрицательных слов [15]; для распознавания лицевой экспрессии счастья его выраженность на лице должна быть более яркой [19].
В большинстве исследований под генетическим базисом распознавания лицевой экс- прессии эмоций подразумевают гены, ассоциированные со структурно-функциональными особенностями фузиформной извилины. Высокий коэффициент наследования имеют как успешное распознавание эмоций, так и лицевая агнозия [8; 11]. Такая генетическая пато- логия, как синдром Уильямса, связана с низким уровнем распознавания эмоциональной экспрессии лиц и с низким IQ [16].
Рассмотрим подробнее генетические факторы распознавания выражений лиц. В ис- следовании J.B. Wilmer с соавт. [44] было показано, что коэффициент корреляции оценок распознавания лиц, которые давали монозиготные близнецы, имеет значение 0,7, в то вре- мя как коэффициент корреляции оценок, которые давали дизиготные близнецы, равнялся 0,29, что свидетельствует о высокой наследуемости данного признака. Подобные резуль- таты были получены в исследовании, проведенном N.G. Shakeshaft и R. Plomin на выборке 2000 близнецов [38]. В исследовании J.Y. Lau с соавт. было также показано значительное влияние наследственных факторов на успешность распознавания лицевых экспрессий [25].
Q. Zhu с соавт. был выявлен высокий коэффициент наследования успешности распоз- навания лиц в группах европейцев и китайцев [47]. Интересно отметить тот факт, что вы- соко наследуется распознавание вертикально ориентированных лиц, но не перевернутых, разделенных на части изображений. Также не получено данных о генетических основаниях распознавания изображений домов, не выявлено связей между общими познавательными способностями и успешностью распознавания лиц. McKone E., Palermo R. заявляют о пре- обладании вклада генетических и эволюционных факторов в распознавание лиц, по сравне- нию со средовыми [31].
В последние годы более пристальное внимание уделяется изучению молекулярно- генетических коррелятов успешности распознавания лицевых экспрессий. Традиционно с навыками распознавания эмоций и контекста социальных ситуаций связывают ген COMT фермента катехол-О-метилтрансферазы, расщепляющей катехоламиновые нейромедиа- торы — дофамин, адреналин и норадреналин [42]. В исследованиях M.N. Smolka с соавт.,
E.M. Drabant с соавт. было показано, что ген COMT связан с выраженным нейронным от- ветом на неприятные стимулы, что проявлялось в активации различных структур лимбиче- ской системы и префронтальной коры, а также способствовало низкой устойчивости носи- телей аллеля Met к негативному настроению [12; 40]. В более ранних исследованиях нами не было выявлено связи между длительностью пребывания моноаминов в синаптической щели (генотипами гена COMT) и временем распознавания эмоций, однако было показано, что полиморфизм Val158Met ассоциирован с точностью распознавания лицевых экспрес- сий и с уровнем эмоционального интеллекта [2].
B. Zhu с соавт. была исследована связь генов дофаминовой системы (синтеза, транс- порта, рецепторов и модуляции) с распознаванием мимики на выборке здоровых китайских студентов [46]. Были получены связи 15 SNP с распознаванием мимических проявлений отвращения, 13 SNP — с распознаванием удивления, 5 SNP— с распознаванием грусти,
1 SNP — с распознаванием страха. Авторы пришли к выводу о том, что значительный вклад в индивидуальные различия в распознавании мимики отвращения вносят генетические ва- риации в подсистемах синтеза и модуляции дофамина. В проведенном нами ранее иссле- довании с более высоким уровнем эмоционального интеллекта был ассоциирован генотип C/С гена дофаминового рецептора второго типа DRD2 [2].
В качестве генов-кандидатов распознавания лицевых экспрессий рассматриваются также гены серотонинергической системы, в число которых входит ген рецептора серотони- на второго типа 5HT2A (HTR2A). Изменения морфофункциональных характеристик ре- цепторов серотонина (5HT) 1A и 2A связывают с дисфункциональными искажениями эмо- циональной обработки при аффективных расстройствах. В исследовании M. Kometer с со- авт. показано, что активация серотониновых рецепторов типов 1A и 2A играет важную роль в регуляции настроения и в распознавании эмоциональных экспрессий здоровыми людьми [23]. M. Matsunaga с соавт. было показано, что аллель G гена HTR2A rs6311 ассоциирован в группах представителей восточных и западных культур с эмоциональным заражением [30]. В исследовании H. Guo с соавт. показано, что генотип C/C гена 5HT2A, а также вы- сокоактивный генотип гена MAOA (ген фермента моноаминоксидазы, расщепляющей мо- ноамины, в том числе серотонин и дофамин) ассоциированы с анормальной активацией в правой средней фронтальной извилине у пациентов с депрессией при распознавании негативных эмоций [17].
Широко обсуждаемой является связь индивидуальных различий в распознавании лиц с геном рецептора окситоцина OXTR, строение которого, в свою очередь, также связы- вают с социальной тревожностью и синдромом Аспергера. В исследовании D.H. Skuse et al. она была показана при помощи теста распознавания лиц Уоррингтон [39], но в исследова- нии R.J. Verhallen с соавт. [41] эта связь не была обнаружена, ни один из исследовавшихся 75 SNP не показал значимых результатов.
Таким образом, гены серотониновой и дофаминовой нейромедиаторных систем мо- гут рассматриваться в качестве генов-кандидатов успешности распознавания лицевых экс- прессий.
В целом, на сегодняшний день на выборке здоровых участников проведено недоста- точно исследований, посвященных изучению генетических факторов распознавания эмо- ций, для понимания фундаментальных механизмов, лежащих в их основе, что и определило цель и задачи настоящего исследования.
Целью данного исследования явилось изучение успешности распознавания лицевых экспрессий носителями различных генотипов генов COMT, DRD4, 5HT2A, MAOA.
Гипотезы исследования:
-
носители разных генотипов генов COMT, DRD4, 5HT2A, MAOA могут отличаться друг от друга по степени успешности распознавания лицевых экспрессий;
-
разная концентрация и продолжительность пребывания нейромедиаторов в синап- тическом пространстве, разная плотность рецепторов дофамина и серотонина, ассоцииро- ванные с различными генотипами исследуемых нами генов, могут быть связаны с различ- ной успешностью распознавания лицевых экспрессий.
Материалы и методыУчастники исследования. В исследовании приняли участие 87 студентов универси- тета (европейцы, женщин — 63%, средний возраст — 20,44 ± 2,6 лет). Все участники имели нормальное или скорректированное до нормы зрение и не имели в анамнезе неврологиче- ских и психических заболеваний. Все участники были информированы о предстоящей про- цедуре и дали письменное согласие на добровольное участие в эксперименте. Исследование было одобрено местной комиссией по этике и выполнялось с соблюдением этических норм в соответствии с этическим кодексом Всемирной медицинской ассоциации (декларация Хельсинки).
Стимулы. В качестве стимулов было использовано 210 фотографий лиц взрослых европейцев анфас, которые были взяты из свободно распространяемых баз данных: MMI [34], KDEF [28], Rafd [24] и WSEFEP [33]. Количество мужских и женских лиц было оди- наковым. Стимулы отображали 6 базовых эмоций [14], а также нейтральное выражение лица. Изображения лиц выравнивались по средней яркости и RMS-контрасту и вписыва- лись в условную окружность диаметром 880 пикселей (22,8 угловых градуса).Процедура. Наблюдатели располагались так, чтобы их глаза находились на расстоя- нии 60 см от центра экрана. Изображения мужских и женских лиц с разным выражением предъявлялись в случайной последовательности на 700 мс. Участник исследования должен был определить выражение показанного лица и сообщить о своем решении нажатием кла- виши мыши на рейтинговой шкале с названиями эмоций (рис. 1). Демонстрация очередно- го стимула запускалась через 100 мс после ответа участника.

Рис. 1. Интерфейс ввода ответа, организованный при помощи компонента RatingScale из библиотеки PsychoPy [35]. Треугольный движок следует за указателем мыши, выбор фиксируется нажатием левой клавиши
Генетический анализ проводился на базе лаборатории медицинской генетики Медико- генетического центра клиники ГБОУ ВПО РостГМУ Минздрава России (г. Ростов-на- Дону). Выделение ДНК осуществлялось из клеток буккального эпителия, затем проводи- лась процедура ПЦР в реальном времени для определения генотипов генов COMT, 5HT2A, DRD4 (амплификатор Real-Time CFX96 Touch, Bio-Rad, США), а также процедура элек- трофореза в агарозном геле для определения числа тандемных повторов промоторного участка MAOA.
Были проанализированы следующие участки ДНК: полиморфный локус Val158Met гена COMT (472A>G, rs4680), полиморфный локус rs6313 гена рецептора серотонина 5HT2A (HTR2A) и полиморфный локус rs1800955 гена рецептора дофамина подтипа D4 (DRD4), тандемные повторы (VNTR) в промоторной области гена моноаминоксидазы А MAOA.
Распределение генотипов изучаемых генов соответствовало равновесию Харди— Вайнберга [29].
Ген МАОА (VNTR): генотипов H (4 повтора промоторного участка гена) — 23 (26%); генотипов L (5 повторов промоторного участка гена) — 42(48%); генотипов M (5/4 повто- ров промоторного участка гена) — 21 (26%).
Ген COMT (Val158Met, rs4680): Val/Val — 11 (13%); Val/Met — 50 (57%); Met/Met—26 (30%).
Ген DRD4 (C-521T, rs1800955): TT — 32 (37%); CT — 40(46%); CC — 15 (17%).
Ген5HT2A (Т102С, rs6313, Tr3): СС— 29 (33%); CT — 46 (53%); TT — 12 (14%).
Обработка результатов. Для каждой эмоции было определено количество пра- вильных узнаваний и «ложных тревог», после чего рассчитан показатель чувствительности (sensitivity index) d’. Для расчёта d’ использовалась функция PAL_SDT_1AFC_PHFtoDP из Palamedestoolbox [21]. В качестве pH рассматривалась вероятность правильной иденти- фикации эмоции (hits), в качестве pF — вероятность ложной (fails). Например, для эмоции «радость» pH равен отношению числа ответов «радость» на стимулы, выражающие радость, к их общему числу; pF же — это отношение числа ответов «радость» на стимулы, не вы- ражающие радость, к общему числу таких стимулов. Фактически функция вычисляла раз- ность нормализованных pH и pF.В случае, если pH оказывался равным 1 (например, все стимулы, выражающие радость, опознаны правильно), pH уменьшался на малую величи- ну 2,2204E-16 («машинный эпсилон»); аналогичным образом увеличивалось значение pF, если оно оказывалось равным нулю (ни один стимул, не выражающий радость, не опознан как радость). Использование данного вычислительного приема позволяет сохранить по- рядковые отношения между вычисленными значениями d’, когда они достаточно велики, по сравнению с заменой 1 или 0 на ближайшее дискретное значение вероятности (напри- мер, 1 на 1/30 или 0 на 1/180).
Для межгрупповых сравнений показателей чувствительности к разным эмоциям но- сителей разных генотипов изучаемых нами генов использовался тест Краскела—Уоллеса. В качестве постхок процедуры применялся тест Данна с поправкой Холма на множествен- ные сравнения [13]. Статистическая обработка полученных результатов осуществлялась с применением свободно распространяемого программного пакета JASP Computer software (Version 0.16, 2021) и программной среды R.
Результаты
Ген COMT
Лицевые экспрессии радости (H=1,5, df=2, p=0,5) и отвращения (H=2,1, df=2, p=0,4) распознавались одинаково успешно носителями разных генотипов гена COMT. Также не было получено достоверных различий в распознавании нейтральных лиц (H=2,9, df=2, p=0,2).
В ходе применения теста Краскела—Уоллеса были получены значимые различия в успешности распознавания лицевой экспрессии удивления (H=7,7, df=2, p=0,02). В ходе апостериорного анализа Данна было получено, что лучше всего данную эмоцию распозна-
ют носители генотипа Val/Val (VM — VV, z=-2,596, pholm=0,014; рис. 2).
Также носители генотипа Val/Val достоверно лучше распознают эмоции печали (H=11,2, df=2, p=0,004; MM — VV, z=-2,667, pholm=0,008, VM — VV, z=-3,336; pholm=0,001;
В ходе апостериорного анализа была выявлена тенденция, не достигающая уров- ня статистической значимости при применении поправки на множественные сравнения, к менее успешному распознаванию эмоции радости носителями генотипа CC гена DRD4 (H=3,3, df=2, p=0,2; CC—CT, z=-1,816, p=0,035, pholm=0,104; рис.7); а также к более успешно- му распознаванию эмоции гнева носителями генотипа CT гена DRD4 (H=4,0, df=2, p=0,1; CT-TT, z=1,988, p=0,023, pholm=0,07; рис.8).
Ген 5HT2A(HTR2A)
У носителей разных генотипов гена 5HT2A не было получено достоверных разли- чий в успешности распознавания эмоций: печали (H=0,999, df=2, p=0,6), страха (H=0,3, df=2, p=0,9), гнева (H=0,2, df=2, p=0,9), отвращения (H=1,2, df=2, p=0,6), удивления (H=1,1, df=2, p=0,6), а также при распознавании нейтральных выражений лиц (H=0,3, df=2, p=0,9).
В ходе апостериорного анализа была выявлена тенденция, не достигающая уров- ня статистической значимости при применении поправки на множественные сравнения, к более успешному распознаванию эмоции радости носителями генотипа TTгена 5HT2A (H=3,9, df=2, p=0,1; CT-TT, z=-1,965, p=0,025, pholm=0,074; рис. 9).
Ген MAOA
Не было выявлено связи между генотипами гена MAOA и успешностью распознава- ния эмоций: удивления (H=0,5, df=2, p=0,8), страха (H=0,7, df=2, p=0,7), гнева (H=2,1, df=2, p=0,4), отвращения (H=1,6, df=2, p=0,5), печали (H=1,4, df=2, p=0,5), радости (H=1,5, df=2, p=0,5), а также при распознавании нейтральных выражений лиц (H=1,8, df=2, p=0,4).
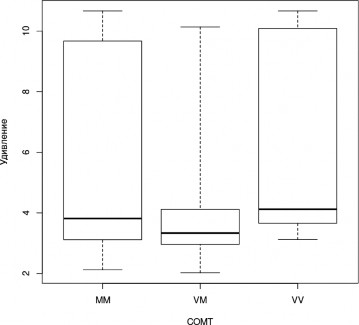
Рис. 2. Успешность распознавания лицевой экспрессии эмоции удивления носителями разных генотипов гена COMT (Kruskal-Wallis Test, p<0,05): «ящик» ограничивает квартили 2 и 3, концы «усов» соответствуют границам диапазона наблюдаемых значений, жирная черта — медиане
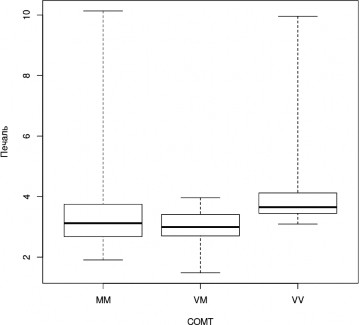
Рис. 3. Успешность распознавания лицевой экспрессии эмоции печали носителями разных генотипов гена COMT (Kruskal-Wallis Test, p<0,05) (условные обозначения — как в рис
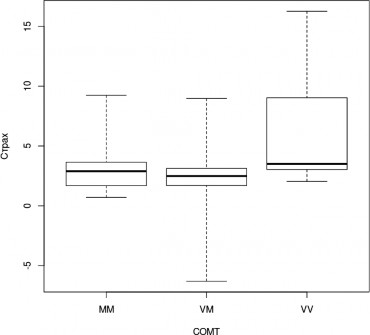
Рис. 4. Успешность распознавания лицевой экспрессии эмоции страха носителями разных генотипов гена COMT, (Kruskal-WallisTest, p<0,05) (условные обозначения — как в рис. 2)
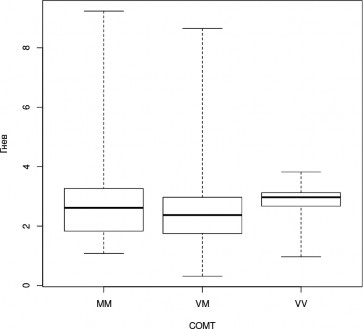
Рис. 5. Успешность распознавания лицевой экспрессии эмоции гнева носителями разных генотипов гена COMT (Kruskal-Wallis Test, p<0,05); (условные обозначения — как в рис. 2)
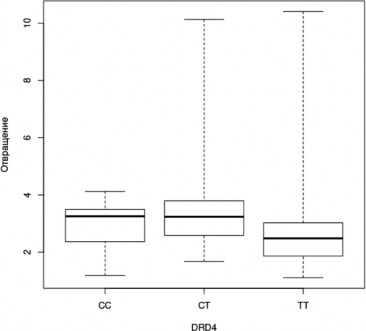
Рис. 6. Успешность распознавания лицевой экспрессии эмоции отвращения носителями разных генотипов гена DRD4 (Kruskal-Wallis Test, p<0,05); (условные обозначения — как в рис. 2)
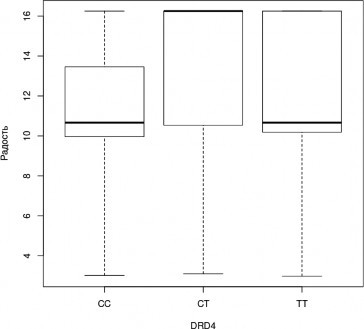
Рис. 7. Успешность распознавания лицевой экспрессии эмоции радости носителями разных генотипов гена DRD4 (Kruskal-Wallis Test, p<0,05); (условные обозначения — как в рис. 2)
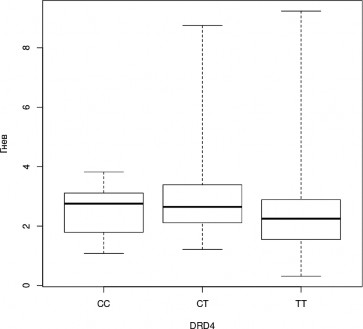
Рис. 8. Успешность распознавания лицевой экспрессии эмоции гнева носителями разных генотипов гена DRD4 (Kruskal-Wallis Test, p<0,05); (условные обозначения — как в рис. 2)
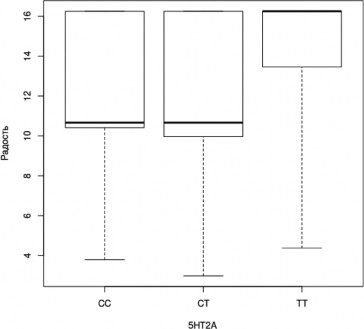
Рис. 9. Успешность распознавания лицевой экспрессии эмоции радости носителями разных генотипов гена 5HT2A (Kruskal-Wallis Test, p<0,05); (условные обозначения — как в рис. 2)
Обсуждение результатов
Полученные нами данные о связи генотипа Val/Val гена COMT c большей успешно- стью распознавания лицевой экспрессии гнева согласуются с результатами исследований других авторов: в работе E. Weiss с соавт. [42], выполненной на выборке из 100 участников, было показано, что гомозиготы Val, по сравнению с гомозиготами Met, лучше и быстрее распознают эмоции гнева и печали.
В нашем исследовании получено, что экспрессия рецептора D4 ассоциирована с особен- ностями восприятия эмоциональных выражений лиц: генотип CT гена DRD4 связан с более успешным распознаванием лицевой экспрессии отвращения, а генотип CC гена DRD4 ассоци- ирован с тенденцией к менее успешному распознаванию эмоции радости. Рецепторы D4 распо- ложены в миндалине, префронтальной коре головного мозга, гиппокампе, переднем таламусе и гипоталамусе, что подчеркивает их связь с эмоциональными процессами [37]. Наши данные согласуются с результатами J.I. Kang с соавт. [20], в исследовании которых была показана связь между геном DRD4 и распознаванием эмоции отвращения. Авторами было показано, что группа без двух повторов аллелей полиморфизма вариабельного числа тандемных повторов (VNTR) лучше распознавала мимическое выражение отвращения, по сравнению с группой носителей двух повторов аллелей. В то же время в исследовании B. Zhu[46], в рамках которого из- учались локусы VNTR, rs12720366 гена DRD4, данная связь не была выявлена. В исследовании
Liu с соавт. [26] была изучена связь генов дофаминергического пути (DBH rs1611115, DDC rs6592961, DRD1 rs251937, DRD2 rs4630328, DRD3 rs167771) с точностью распознавания эмо- ций у детей с РАС. Было показано, что дефицит распознавания эмоций на лице связан с не- которыми SNP генов дофаминергического пути: точность распознавания страха была связана с экспрессией рецептора D1 (DRD1 rs251937); точность распознавания печали была связана с экспрессией рецептора D2 (DRD2 rs4630328); экспрессия рецептора D3 (DRD3 rs167771) свя- зана со временем распознавания радости, печали и страха, с точностью распознавания гнева и страха, а также с риском развития расстройств аутистического спектра.
Ранее нами было получено, что носители генотипа A/A гена HTR2A (5HT2A) имеют более высокие показатели эмоционального интеллекта, по сравнению с другими группами [2]. Данный результат согласуется с результатом, полученным в ходе настоящего исследо- вания: носители генотипа TT гена 5HT2A более успешно (на уровне тенденции) распознают эмоцию радости (локусы tr2 —генотипы AA, AG, GG и tr3 —генотипы TT, TC, CC данного гена находятся в сцеплении). В то же время, в исследовании M. Matsunaga с соавт. показано, что носители аллеля G имеют более высокие показатели эмоционального заражения [30].
В данном исследовании нами не было обнаружено результатов относительно успешности дифференциации эмоциональных выражений лиц носителями разных генотипов гена МАОА, однако в более ранних исследованиях нами было получено, что женщины с высокоактивным и гетерозиготным генотипами МАОА более успешно справляются с задачами дискриминации и оценки эмоционально окрашенных стимулов (сцен) [3]. В исследовании B. Zhu [45], проведенном на выборке здоровых китайских студентов, при вычислении основных эффектов была выявлена связь между полиморфизмом числа тандемных повторов гена MAOA VNTR и распознаванием лицевых экспрессий отвращения и удивления; при проведении множественного регрессионно- го анализа, направленного на изучение общего вклада основных эффектов для локусов дофами- нергической системы, эти результаты не были подтверждены. Неоднозначные результаты могут быть связаны с недостаточной численностью выборки, а также со спецификой эмоционально окрашенных стимулов — механизмы распознавания лиц и сцен могут различаться.
Выводы
Носители разных генотипов генов COMT, DRD4, 5HT2A (HTR2A), MAOA отлича- ются друг от друга по степени успешности распознавания лицевых экспрессий.
С быстрым обратным захватом катехоламинов пресинаптической мембраной (гено- тип Val/Val гена COMT) связаны достоверно лучшее распознавание эмоций удивления, страха, печали, а также тенденция к лучшему распознаванию эмоции гнева.
Средний уровень экспрессии рецептора D4 (генотип C/T гена DRD4) связан с более успешным распознаванием лицевой экспрессии отвращения.
Высокий уровень экспрессии рецептора D4 (генотип C/C гена DRD4) ассоциирован с менее успешным распознаванием эмоции радости (на уровне тенденции).
Высокая плотность рецепторов серотонина второго типа на постсинаптической мем- бране нейрона (генотип T/T гена 5HT2A (HTR2A) ассоциирована с более успешным рас- познаванием эмоции радости (на уровне тенденции).
Не обнаружено связи между генотипами гена MAOA и успешностью распознавания эмоций.
Ограничения исследования
В данной работе представлены результаты, полученные на
выборке из 87 человек, что может быть причиной недостаточно выраженных
статистических эффектов, полученных для генов 5HT2A (HTR2A), COMT и DRD4,
которые мы описали на уровне тенденций. Увеличение объема выборки позволит
уточнить полученный на данном этапе результат.
рис.3) и страха (H=10,5, df=2, p=0,005; MM — VV, z=-2,123, pholm=0,03, VM — VV, z=-3,199, pholm=0,002; рис. 4).
В ходе пост-хок анализа Данна была обнаружена тенденция, не достигающая уровня статистической значимости при применении поправки на множественные сравнения, к более успешному распознаванию эмоции гнева носителями генотипа Val/Val по сравнению с носи- телями генотипа Val/Met (H=3,0, df=2, p=0,2; VM — VV, z=-3,336, p=0,046, pholm=0,1; рис. 5).
Литература
-
Беловол Е.В., Хворова Е.М. Когнитивный стиль личности как фактор эффективного распознавания лицевой экспрессии // Вестник Российского университета дружбы народов. Серия: Психология и педагогика. 2015. № 3. С. 51—60.
-
Воробьева Е.В., Косоногов В.В., Ковш Е.М. Эмоциональный интеллект: генетические и психофизиологические корреляты. М.: Мир науки, 2021. 96 с.
-
Ермаков П.Н., Ковш Е.М., Воробьева Е.В. Особенности вызванной активности мозга девушек- носительниц различных генотипов по маркеру MAOA-uVNTR при оценке эмоционально окрашенных сцен [Электронный ресурс] // Российский психологический журнал. 2016. Том 13. № 4. С. 232—253. DOI:10.21702/rpj.2016.4.14
-
Карабущенко Н.Б., Хворова Е.М. Социально-интеллектуальные особенности распознавания лицевой экспрессии //Лицо человека: познание, общение, деятельность:коллективная монография / Отв. ред. К.И. Ананьева, В.А. Барабанщиков, А.А. Демидов. М.: Когито-Центр; Московский институт психоанализа, 2019. С. 395—407.
-
Культурно-интеллектуальные особенности распознавания лицевой экспрессии представителей других этносов / Н.Б. Карабущенко [и др.] // Российский психологический журнал. 2017. Том 14. № 3. С. 169—189. DOI:10.21702/rpj.2017.3.9
-
Лунякова Е.Г., Райков А.В., Махкамова Д.З. Проявление механизмов целостного восприятия в распознавании лицевой экспрессии у испытуемых разных возрастных групп // Лицо человека в контекстах природы, технологий и культуры: коллективная монография / Отв. ред. К.И. Ананьева, В.А. Барабанщиков. М.: Когито-Центр; Московский институт психоанализа, 2020. С. 194—209.
-
Babenko V. et al. Relationship Between Facial Areas With the Greatest Increase in Non-local Contrast and Gaze Fixations in Recognizing Emotional Expressions // International Journal of Cognitive Research in Science, Engineering and Education. 2021. Vol. 9. № 3. P. 359—368. DOI:10.23947/2334-8496-2021-9- 3-359-368
-
Bowles D. et al. Diagnosing prosopagnosia: Effects of ageing, sex, and participant—stimulus ethnic match on the Cambridge Face Memory Test and Cambridge Face Perception Test // Cognitive neuropsychology. 2009. Vol. 26. № 5. P. 423—455. DOI:10.1080/02643290903343149
-
Chen D., Song P. Dual-graph regularized discriminative transfer sparse coding for facial expression recognition // Digital Signal Processing. 2021. Vol. 108. 102906. DOI:10.1016/j.dsp.2020.102906
-
Davis J.M. et al. Individual differences in the ability to recognize facial identity are associated with social anxiety // PLoS ONE. 2011. Vol. 6. № 12. Article e28800. DOI:10.1371/journal.pone.0028800
-
De Haan E. A familial factor in the development of face recognition deficits // Journal of Clinical and Experimental Neuropsychology. 1999. Vol. 21. P. 312—315. DOI:10.1076/jcen.21.3.312.917
-
Drabant E.M. et al. Catechol-O-methyltransferase val158met genotype and neural mechanisms related to affective arousal and regulation // Archives of general psychiatry. 2006. Vol. 63. № 12. P. 1396—1406. DOI:10.1001/archpsyc.63.12.1396
-
Dunn O.J. Multiple Comparisons Using Rank Sums // Technometrics. Taylor & Francis. 1964. Vol. 6. № 3. P. 241—252. DOI:10.1080/00401706.1964.10490181
-
Ekman P. An argument for basic emotions // Cognition & emotion. 1992. Vol. 6. № 3-4. P. 169—200. DOI:10.1080/02699939208411068
-
Erickson K. et al. Mood-congruent bias in affective go/no-go performance of unmedicated patients with major depressive disorder // American Journal of Psychiatry. 2005. Vol. 162. № 11. P. 2171—2173. DOI:10.1176/appi.ajp.162.11.2171
-
Gagliardi C. et al. Facial expression recognition in Williams syndrome// Neuropsychologia. 2003. Vol. 41. № 6. P. 733—738. DOI:10.1016/S0028-3932(02)00178-1
-
Guo H. et al. Synergistic effect of 5 HT2A receptor gene and MAOA gene on the negative emotion of patients with depression // Clinical physiology and functional imaging. 2014. Vol. 34. № 4. P. 277—281. DOI:10.1111/cpf.12094
-
Jamieson A.J., Davey C.G., Harrison B.J. Differential Modulation of Effective Connectivity in the Right Hemisphere of the Brain’s Extended Face Processing System by Fearful and Sad Facial Expressions // Eneuro. 2021. Vol. 0380. №. 20. DOI:10.1523/ENEURO.0380-20.2021
-
Joormann J., Gotlib I. H. Is this happiness I see? Biases in the identification of emotional facial expressions in depression and social phobia // Journal of abnormal psychology. 2006. Vol. 115. № 4. P. 705. DOI:10.1037/0021-843X.115.4.705
-
Kang J.I. et al. Association of DRD4 and COMT polymorphisms with disgust sensitivity in healthy volunteers // Neuropsychobiology. 2010. Vol. 61. № 2. P. 105—112. DOI:10.1159/000275822
-
Kingdom F.A.A., Prins N. Psychophysics: A practical introduction (Second edition). Amsterdam: Elsevier/Academic Press, 2016. 331 p. DOI:10.1016/C2012-0-01278-1
-
Kohn N. et al. Cognitive Control of Emotions in Challenging Contexts // Frontiers in Behavioral Neuroscience. 2021. Vol. 15. 785875. DOI:10.3389/fnbeh.2021.785875
-
Kometer M. et al. Psilocybin biases facial recognition, goal-directed behavior, and mood state toward positive relative to negative emotions through different serotonergic subreceptors// Biological psychiatry. 2012. Vol. 72. № 11. P. 898—906. DOI:10.1016/j.biopsych.2012.04.005
-
Langner O. et al. Presentation and validation of the Radboud Faces Database // Cognition and emotion. 2010. Vol. 24. № 8. P. 1377—1388. DOI:10.1080/02699930903485076
-
Lau J.Y. et al. Individual differences in children’s facial expression recognition ability: The role of nature and nurture // Developmental neuropsychology. 2009. Vol. 34. № 1. P. 37—51. DOI:10.1080/87565640802564424
-
Liu Z. et al. Facial Emotion Recognition and Polymorphisms of Dopaminergic Pathway Genes in Children with ASD // Behavioural neurology. 2020. 6376842. DOI:10.1155/2020/6376842
-
Lopatina O.L. et al. Neurobiological aspects of face recognition: the role of oxytocin // Frontiers in behavioral neuroscience. 2018. Vol. 12. № 195. DOI:10.3389/fnbeh.2018.00195
-
Lundqvist D., Flykt A., Öhman A. The Karolinska directed emotional faces (KDEF). [Database record]. APA PsycTests. KarolinskaInstitutet. 1998. DOI:10.1037/t27732-000
-
Mayo O. A Century of Hardy—Weinberg Equilibrium // Twin Research and Human Genetics.2008. Vol. 11. № 3. P. 249—256. DOI:10.1375/twin.11.3.249
-
Matsunaga M. et al. Serotonin Receptor (HTR2A) Gene Polymorphism Modulates Social Sharing of Happiness in Both American and Japanese Adults // Japanese Psychological Research. 2022. Vol. 64. № 2. P. 181—192. DOI:10.1111/jpr.12389
-
McKone E., Palermo R. A strong role for nature in face recognition // Proceedings of the National Academy of Sciences. 2010. Vol. 107. № 11. P. 4795—4796. DOI:10.1073/pnas.1000567107
-
Nelson C., De Haan M. A neurobehavioral approach to the recognition of facial expressions in infancy // The Psychology of Facial Expression (Studies in Emotion and Social Interaction)/ G. Mandler, J. Russell,
J. Fernández-Dols (Eds.). Cambridge: Cambridge University Press, 1997. P. 176—204. DOI:10.1017/ CBO9780511659911.010
-
Olszanowski M. et al. Warsaw set of emotional facial expression pictures: a validation study of facial display photographs // Frontiers in psychology. 2015. Vol. 5.51516.DOI:10.3389/fpsyg.2014.01516
-
Pantic M. et al. Web-based database for facial expression analysis. Proceedings of the IEEE International Conference on Multimedia and Expo (Amsterdam, the 25 of October, 2005). Amsterdam. IEEE. 2005. 5 p. DOI:10.1109/ICME.2005.1521424
-
Peirce J. et al. PsychoPy2: Experiments in behavior made easy // Behav Res Methods. 2019. Vol. 51. № 1. P. 195—203. DOI:10.3758/s13428-018-01193-y
-
PrinsN. Applying the model comparison approach to tests pecific research hypotheses in psychophysical research using the Palamedes toolbox // Frontiers in psychology. 2018. Vol. 9. 1250. DOI:10.3389/fpsyg.2018.01250
-
Salgado-Pineda P. et al. Dopaminergic contribution to the regulation of emotional perception // Clinical neuropharmacology. 2005. Vol. 28. № 5. P. 228—237. DOI:10.1097/01.wnf.0000185824.57690.f0
-
Shakeshaft N.G., Plomin R. Genetic specificity of face recognition // Proceedings of the National Academy of Sciences. 2015. Vol. 112. № 41. P. 12887—12892. DOI:10.1073/pnas.1421881112
-
Skuse D.H. Common polymorphism in the oxytocin receptor gene (OXTR) is associated with human social recognition skills // Proceedings of the National Academy of Sciences. USA. 2014. Vol. 111. P. 1987— 1992. DOI:10.1073/pnas.1302985111
-
Smolka M.N., et al. Catechol-O-methyltransferase val158met genotype affects processing of emotional stimuli in the amygdala and prefrontal cortex //Journal of Neuroscience. 2005. Vol. 25. №4. P. 836—842. DOI:10.1523/JNEUROSCI.1792-04.2005
-
Verhallen R.J. et al. The oxytocin receptor gene (OXTR) and face recognition // Psychological science. 2017. Vol. 28. № 1. P. 47—55. DOI:10.1177/0956797616672269
-
Weiss E. et al. Differential effect of catechol-O-methyltransferase Val158Met genotype on emotional recognition abilities in healthy men and women // Journal of the International Neuropsychological Society. 2007. Vol. 13. № 5. P. 881—887. DOI:10.1017/S1355617707070932
-
Wilmer J.B. Individual differences in face recognition: A decade of discovery // Current Directions in Psychological Science. 2017. Vol. 26. № 3. P. 225—230. DOI:10.1177/0963721417710693
-
Wilmer J.B. et al. Human face recognition ability is specific and highly heritable // Proceedings of the National Academy of sciences. 2010. Vol. 107. № 11. P. 5238—5241. DOI:10.1073/pnas.0913053107
-
World Medical Association. Code of Ethics of the World Medical Association: Declaration of Helsinki (ed.). Helsinki: WMA.1964. 177 p.
-
Zhu B. et al. Genetic variations in the dopamine system and facial expression recognition in healthy Chinese college students // Neuropsychobiology. 2012. Vol. 65. № 2. P. 83—89. DOI:10.1159/000329555
-
Zhu Q. et al. Heritability of the specific cognitive ability of face perception // Current Biology. 2010. Vol. 20. № 2. P. 137—142. DOI:10.1016/j.cub.2009.11.067
Информация об авторах
Метрики
Просмотров
Всего: 450
В прошлом месяце: 23
В текущем месяце: 0
Скачиваний
Всего: 115
В прошлом месяце: 4
В текущем месяце: 0