Марютина Т.М., доктор психологических наук, профессор, зав. кафедрой дифференциальной психологии и психофизиологии института психологии имени Л.С. Выготского, РГГУ, Москва, Россия, t.m.mariutina@rambler.ru
«Исчезающая наследуемость» - лингвистическая метафора, отражающая феномен рассогласования оценок влияния генотипа на функционирование психики в норме и патологии, которые фиксируются двумя разными методами: популяционным и молекулярно-генетическим. С одной стороны, в популяционных исследованиях лиц, состоящих в родстве (близнецы, сиблинги, родители-дети), при оценке их когнитивных и личностных особенностей были получены достаточно высокие значения основного статистического показателя - наследуемости. С другой стороны, в молекулярно-генетических исследованиях было установлено, что отдельные генетические полиморфизмы не могут объяснить большую часть наследуемости такого рода особенностей психики как в норме, так и при психических расстройствах. В статье рассматривается история вопроса, анализируются возможные причины возникновения данного феномена и пути выхода из сложившейся ситуации.
Введение
Словосочетание «старая проблема» в заглавии данной статьи подразумевает проблему соотношения генетического и средового в формировании индивидуально-психологических признаков человека в норме и патологии [Lartseva, 2014; Maryutina, 2013; R. Plomin, et, 2016]. В ХХ в. был проведен целый ряд исследований, главным предметом изучения в которых служили показатели наследуемости этих признаков. По определению, наследуемость (heritability) - доля фенотипической изменчивости в популяции, обусловленная генетической изменчивостью (в отношении к определенному качественному или количественному признаку). Это статистический показатель, имеющий количественное выражение и получаемый эмпирическим путем при исследовании популяций (реально больших выборок) лиц, являющихся родственниками, т. е. имеющими частично общие гены. Наследуемость не следует путать с понятием «наследственность» (heredity), представляющим способность живых организмов передавать потомкам морфологические, физиологические и этологические признаки.
В ходе популяционных исследований, опирающихся на эмпирически получаемые показатели, был получен большой объем конкретных материалов. Эти работы подтвердили важную роль генетических факторов в межиндивидуальной вариативности общего интеллекта, отдельных способностей, темперамента, личности. Было установлено, что многие из психических расстройств также имеют наследственную основу [Grigorenko, 2010; Lartseva, 2014; Maryutina, 2013; Goldsmith, 1984; Lee, 2011]. Однако в последние годы на этом относительно уравновешенном фоне появились статьи с парадоксальным словосочетанием «исчезающая наследуемость» (missing heritability) [Manolio, 2009; Visvikis-Siest, 2013; Zhu, 2015; Zuk, 2013; Zuk, 2011]. Цель настоящей статьи — проанализировать содержание данного термина, выяснить причины и условия его возникновения и предполагаемые способы разрешения возникшего противоречия.
Основная часть
В 2003 г. официально был завершен проект по расшифровке генома человека. Термином «геном» в настоящее время обозначают полный состав ДНК клетки, т. е. совокупность всех генов и сопутствующих им компонентов. Принципы построения геномов и их структурно функциональную организацию изучает новая область знания - геномика. В отличие от традиционных задач психогенетики, задача геномики - идентификация конкретных генов и их роли в определении различных признаков человека, в том числе генов, отвечающих за вариативность высших психических функций в норме и при расстройствах психики [Plomin, 2013].
Различия между этими двумя направлениями исследований роли генотипа и среды в происхождении психологических признаков представлены в табл. 1 [Goldsmith, 1984]. Очевидно, что эти две ветви генетики различаются радикально, начиная от объекта исследования и кончая способами обработки и интерпретации данных. Модели количественной генетики характеризуют наследуемость (качество, которое нельзя оценить применительно к индивидууму), а модели генетики развития адресуются к изучению собственно наследственности как конкретному механизму передачи признаков в череде поколений.
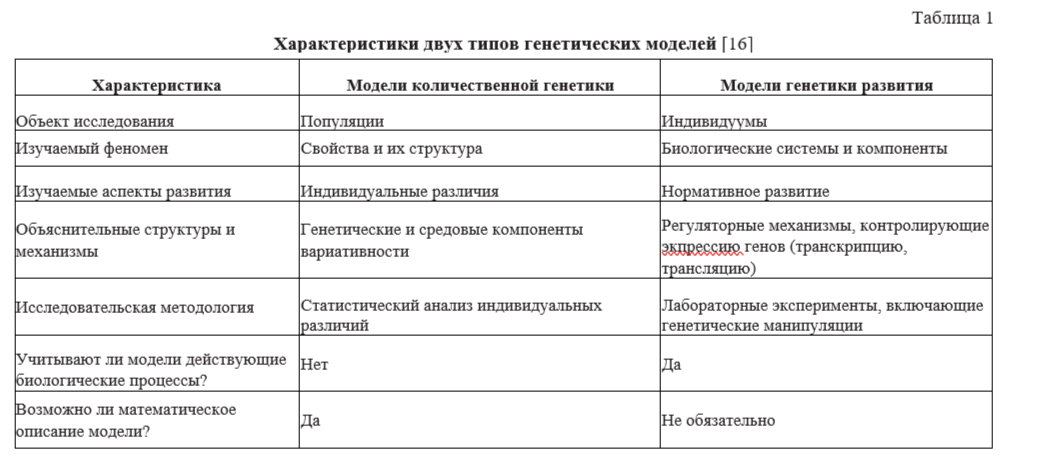
Известно, что подавляющее большинство психологических признаков относится к категории количественных, а именно: измеряемых характеристик, которые различаются у разных индивидуумов по степени выраженности (например, коэффициент интеллекта). Такие фенотипы являются продуктом действия двух или более генов, поэтому их относят к полигенным эффектам. В отличие от моногенных, полигенные признаки не подчиняются законам Менделя. Эти признаки имеют континуально непрерывный характер и обычно варьируют с равномерным отклонением, изображаемым кривой нормального распределения.
Многие наследственные заболевания имеют поли- генную природу; к таковым относятся шизофрения, аутизм, болезнь Альцгеймера и др. [Grigorenko, 2010; Lartseva, 2014; Maryutina, 2013; Lee, 2011]. Большинство фенотипических характеристик в этих случаях являются результатом взаимодействия множества генов и соответствующих локусов количественных признаков, сокращенно - ЛКП (Quantitative Trait Loci - QTLs). Последние являются участками ДНК, либо содержащими гены, либо сцепленными с генами, которые отвечают за тот или иной количественный признак.
Тем не менее, технологический прогресс, достигнутый в ходе реализации проекта «Геном человека», открыл новые возможности в плане идентификации генов, в том числе и для количественных признаков.
Одной из признанных стратегий поиска стало изучение генов-кандидатов, т. е. генов, потенциально ответственных за формирование конкретных признаков. В качестве примера эффективности такого подхода можно привести следующие данные.
Один из наиболее часто упоминаемых в этой связи генов - ген катехол-О-метилтрансферазы СОМТ, фермент, принимающий участие в разрушении медиаторов дофамина. Он играет ключевую роль в регуляции процесса передачи дофамина в префронтальной коре [Alfimova, 2011; Grigorenko, 2010; Barnett, 2008]. Ген расположен на хромосомном участке 22q11. В данном локусе обнаружен ряд полиморфизмов, среди которых Val158Met, ведущий к замене кодируемой аминокислоты, изучается наиболее интенсивно. В случае аллеля Met активность фермента в 3-4 раза ниже, чем в случае аллеля Val. Таким образом, носители аллеля Met предположительно отличаются более высокой активностью дофамина в префронтальной коре за счет его менее эффективного превращения в метаболиты под действием СОМТ [DeYoung, 2011]. Носители аллеля Met, как правило, успешнее выполняют когнитивные задачи, демонстрируют лучшие способности к чтению и пониманию прочитанного [Barnett, 2008]. Была также установлена связь между полиморфизмом гена СОМТ и такой психологической характеристикой, как открытость опыту из пятифакторной модели личности [DeYoung, 2011]. Ген СОМТ далеко не единственный в списке генов-кандидатов, связываемых с психологическими признаками как в норме, так и в патологии [Alfimova, 2011; Grigorenko, 2010; Barnett, 2008; Tunbridge, 2006]. Этот перечень весьма значителен, однако в данном контексте важно другое: в исследованиях генов-кандидатов феномен исчезающей наследуемости не наблюдается.
Данный феномен был обнаружен при применении методов полногеномного анализа генетических ассоциаций сложных признаков (Genomewide association studies - GWAS) [Lartseva, 2014; Maryutina, 2013; Chen, 2015; Lee, 2011]. При таком подходе для поиска локусов, контролирующих признак, используется большое количество маркеров, охватывающих весь геном [Ehret, 2012; Lee, 2011]. Высокая наследуемость некоторых признаков психики и поведения, полученных при исследовании близнецов и приемных детей, дали основания предположить, что этот метод мог бы выявить многочисленные гены, оказывающие существенный эффект на психологические признаки.
Схема такого рода исследований носит стандартный характер. В них обычно сравнивают геномы группы лиц с психологическими особенностями (например, высокий уровень интеллекта или наличие какого- либо заболевания с геномами контрольной группы, т. е. аналогичных по возрасту, полу и другим признакам индивидуумов, не имеющих таких особенностей). Материалом для исследования являются образцы ДНК каждого участника исследования. У них измеряют интересующий параметр, секвенируют ДНК (т. е. считывают последовательность нуклеотидов в ДНК), и затем для каждого полиморфизма подсчитывается вероятность того, что этот полиморфизм статистически значимо влияет на выбранный параметр. Если удается выявить совокупность аллелей, которые значимо чаще встречаются у людей с изучаемым признаком, то говорят, что такой вариант связан, или ассоциирован с ним. Таким образом, в отличие от методов, которые проверяют один или несколько конкретных участков генома, полногеномный поиск ассоциаций использует полную последовательность ДНК.
Предполагалось, что указанный метод позволяет обнаруживать ранее неизвестные гены, имеющие отношение к изучаемому признаку или заболеванию [Alfimova, 2011; Grigorenko, 2010; Maryutina, 2013; Lee, 2011; Manolio, 2009]. По этой причине Психиатрический консорциум полногеномного анализа (Psychiatric GWAS Consortium; PGC: https://pgc.unc.edu/pgc/index. php) поставил своей целью объединение многих исследований с целью идентификации надежных ассоциаций. С помощью GWAS предполагалось также установить связь между количественными признаками, в первую очередь уровнем интеллекта, и определенным полиморфизмом в ДНК [Plomin, 2013; Richardson, 2013]. Первые результаты этих исследований казались обнадеживающими, так что к 1998 г. в некоторых газетах появились сообщения о том, что найдены «гены интеллекта» (например: New York Times. 1998. May 14th) [Richardson, 2013].
Ранние предположения, однако, не нашли подтверждения в последующих исследованиях. В подавляющем большинстве случаев применительно к когнитивным способностям и психическим нарушениям выявленные ассоциации оказались малы, составляя только незначительную часть (<5%) от величины, предполагаемой на основе популяционных исследований наследуемости [Lartseva, 2014; Manolio, 2009; Plomin, 2013; Richardson, 2013].
Аналогичная картина фиксируется в генетической психиатрии [Lartseva, 2014; Maryutina, 2013; Lee, 2011]. Например, аутизм - заболевание с очень высоким вкладом наследственности, предположительно более 90%. Упоминается несколько десятков генов, наиболее значимо связанных с аутизмом. Но если сложить вместе эффект всех этих генов, можно объяснить не более 5% дисперсии. 85% наследуемости не получает адекватного объяснения.
В этом и заключается феномен исчезающей наследуемости. Анализируя его истоки, исследователи указывают на мультифакторную природу данного явления [Lartseva, 2014; Manolio, 2009; Plomin, 2013] и называют в качестве причин следующие факторы.
1. Показатели наследуемости, полученные в исследованиях близнецов и приемных детей, возможно, были завышены. Причина в том, что наследуемость - статистический показатель, который оценивает гено- тип-средовые соотношения в популяции по принципу «здесь и сейчас». Изменения, которые претерпевает популяция (выборка) по своему составу и условиям существования обязательно влекут за собой изменение показателя наследуемости. Так, увеличение разнообразия среды должно привести к снижению наследуемости. Однако, как подчеркивает Р. Пломин [Plomin, 2013], даже если наследуемость была искусственно завышена примерно в два раза и составляет 25% вместо 50%, все еще будет оставаться «зазор», потому что доля известных ассоциаций на сегодняшний день составляет менее 5% дисперсии и для распространенных заболеваний, и для количественных признаков.
2. Недостаточные разрешающие возможности метода полногеномного исследования. Не исключено, что существующая стандартная процедура не дает возможность вскрыть все ресурсы генома, участвующие в детерминации количественных признаков и ряда заболеваний. Допускается ряд усовершенствований метода в двух направлениях: со стороны биохимических основ и со стороны статистической обработки результатов [Lartseva, 2014; Ehret, 2012; Eichler, 2010].
Известно, что в полногеномном анализе широко используются так называемые однонуклеотидные полиморфизмы, далее ОНП (Single nucleotide polymorphism, SNP). Их особенность - отличия последовательности ДНК размером в один нуклеотид (A, T, G или C). Они применяются в генетическом картировании как маркеры с высоким разрешением, благодаря их количеству и стабильной преемственности в ряду поколений. Большое значение имеют ОНП в исследованиях разных групп испытуемых, поскольку их используют для сравнения участков генома между исследуемыми группами (экспериментальной и контрольной). Закономерно, что именно эта технология становится предметом усовершенствования. Модификация метода, предложенная в работе Дж. Эхрета с соавторами [Ehret, 2012], включающая укрупнение единиц анализа в виде построения многопрофильного ОНП на основе линейной комбинации отдельных полиморфизмов, позволяет существенно улучшить результаты, повысить учитываемую долю влияния генов на изучаемые признаки и приблизиться к решению проблемы исчезновения наследуемости. Видимо, исследования в этом направлении будут продолжены.
3. Различия в частоте распространения аллелей. По степени встречаемости в популяциях полиморфные гены и аллели образуют своеобразный континуум, на одном полюсе которого широко распространенные и известные варианты генов и аллелей, а на другом — редкие и, возможно, еще и неустановленные их варианты.
Вклад распространенных вариантов генов приводит к небольшим эффектам. В то же время редко встречающиеся варианты генов могут вносить более существенный вклад в детерминацию количественных признаков или заболеваний. Усилия большинства исследователей в основном сосредоточены на изучении общих генетических вариантов. Высказывается предположение, что какая-то часть исчезающей наследуемости связана с редкими генетическими вариантами [Lee, 2011; Zuk, 2013]. Однако исследование редких образцов представляет собой довольно сложную с методической точки зрения задачу, в первую очередь, в вопросе верификации и воспроизведения результатов и выводов, поэтому предлагается разрабатывать новые подходы к изучению таких вариантов [Zuk, 2011].
Важно также подчеркнуть, что предположение, согласно которому низкочастотные варианты обязательно будут иметь значение в плане генетической детерминации признаков, не имеет под собой оснований. И уже есть исследования (на материале аутоиммунных заболеваний), результаты которых свидетельствуют об отсутствии связи между частотой встречаемости гена и степенью его влияния на признак [Hunt, 2013].
4. Среди других возможных причин исчезновения наследуемости чаще всего упоминается эпистаз (взаимодействие генов, при котором активность одного гена находится под влиянием другого гена, неаллельного ему) [Slatkin, 2009; Trerotola, 2015; Visvikis-Siest, 2013]. Помимо этого, существенное влияние на механизм наследования оказывают условия среды, например, нарушение нормативных требований экологии в период вынашивания плода.
Возможность негенетических наследуемых влияний на фенотип вызывает большой интерес, поскольку выявляет дополнительный механизм наследственной изменчивости, а именно тот, который не обнаруживается в геномных исследованиях. Наследственные эпигенетические изменения были предложены в качестве объяснения исчезающей наследуемости. Однако необходимы молекулярно-генетические данные, чтобы окончательно определить важность эпистаза [Plomin, 2013].
Наряду с этим определенный вклад в обсуждение проблемы вносят и традиционные психогенетические исследования наследуемости. В структуре этого показателя принято выделять две составляющие: аддитивную и доминантную. Первая, определяемая как наследуемость в «узком» смысле слова, характеризует прямую передачу наследственных влияний от родителей к потомкам. Вторая составляющая - доминантная - отражает эффекты взаимодействия генов внутри генома (в том числе и эпистатические). Будучи объединены воедино, они образуют наследуемость в «широком» смысле слова. Предпринимались попытки установить, какой из двух компонентов генетической дисперсии (аддитивный или доминантный) в большей степени обнаруживает себя в исчезновении наследуемости [11;10]. Оказалось, однако, что вклад доминантной дисперсии в общую картину полиморфизмов объясняет лишь небольшую часть фенотипической изменчивости комплексных черт и мало способствует решению проблемы исчезновения наследуемости в узком смысле [Zhu, 2015].
Главный вывод исследований ассоциаций полногеномного анализа состоит в том, что эффект исчезновение наследуемости, вероятно, является результатом участия в генетической детерминации сложных признаков многих ДНК вариантов с небольшим эффектом действия; Возможно, существуют редкие варианты аллелей с большим эффектом действия, но они вряд ли будут главным источником наследуемости в популяции. Два практических следствия вытекают из вывода, что наследственность обусловлена многими ДНК- вариантами небольшого эффекта. Во-первых, будет трудно определить ассоциации ДНК с небольшим размером эффекта и еще труднее будет воспроизвести такие ассоциации. Во-вторых, поскольку варианты ДНК с небольшим эффектом действия установлены, их применение к исследованию развития потребует полигенных исследований, включающих в анализ сотни и тысячи генов [Plomin, 2013; Visscher, 2012].
На этом фоне намечаются и принципиально новые подходы к изучению проблем генетической детерминации сложных признаков [Kolchanov, 2013; Heng, 2011; Zuk, 2013]. Исследования последнего десятилетия свидетельствуют о том, что подавляющее большинство фенотипических признаков человека (биохимических, физиологических, морфологических, поведенческих и т. д.) контролируются очень сложным образом. В основе формирования этих признаков лежат так называемые генные сети, т. е. группы координированно функционирующих генов, взаимодействующих друг с другом как через свои первичные продукты (РНК и белки), так и через разнообразные метаболиты и другие вторичные продукты функционирования самих сетей. Характерное свойство генных сетей, функционирующих в живых системах, состоит в том, что в них регуляторная компонента является ведущей, определяя режим их функционирования и, соответственно, процесс формирования признаков. Это касается как метаболической, так и генетической компоненты генных сетей и проявляется на каждом иерархическом уровне их организации [Kolchanov, 2013].
Хотя общее понимание механизмов функционирования генов на основе законов биологии значительно улучшилось, постепенно становится ясно, что функции генома не могут быть полностью объяснены на основе отдельных генов и их ассоциаций. Гены и геном представляют различные уровни генетической организации со своими собственными системами кодирования; Гены - это только части кода, которые отвечают за белок и РНК, но геном кодирует структуру генетических сетей, которые определяются целым набором генов, хромосом и их топологических взаимодействий в клетке. Соответственно, генетический код ДНК предлагает ограниченное понимание функций генома [Heng, 2011].
С этой точки зрения, предлагается приступить к разработке теории генома, который предусматривает переход от исследований, ориентированных на изучение отдельных генов, к геномным исследованиям. Для того, чтобы совершить подобный переход необходимо признать важность создания новых технологических платформ для декодирования генома и исследования механизмов его функционирования как целого.
Заключение
Понятие «исчезающая наследуемость» фактически фиксирует кризис, возникший при изучении генетической детерминации сложных признаков около 10 лет назад. Суть кризиса в стабильно фиксируемом расхождении результатов оценки генетических влияний на изменчивость количественных признаков и источники происхождения многих заболеваний при изучении их на двух разных уровнях анализа: молекулярно-генетическом и популяционном. Причины расхождения имеют многофакторную структуру. В этот ряд включают: искусственно возникающее превышение популяционных значений наследуемости; недостаточную точность полногеномного анализа, не позволяющую воспроизвести все тонкие механизмы генетической детерминации полигенного признака; преобладание в исследованиях часто встречающихся аллелей, вносящих незначительный вклад в генетическую детерминацию; наличие эпистатиче- ских эффектов и генотип-средового взаимодействия, а также ряд других возможных причин. В связи с вышесказанным рассматриваются новые пути для выхода из кризиса, предполагающие, в первую очередь, совершенствование существующих методов анализа и развитие новых подходов, прежде всего на уровне генома как целого.