Экспериментальная психология
2013. Том 6. № 4. С. 79–95
ISSN: 2072-7593 / 2311-7036 (online)
Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Gryllus assimilis*
Аннотация
Общая информация
* Исследование поддержано грантом Президента Российской Федерации № МК-2816.2012.6.
Ключевые слова: психическое отражение, самоотражение, отражение внешней среды, филогенез психики, насекомые, сверчки
Рубрика издания: Эволюционная и сравнительная психология
Тип материала: научная статья
Для цитаты: Хватов И.А., Харитонов А.Н., Соколов А.Ю. Особенности соотнесения физических характеристик собственного тела с объектами окружающей среды при ориентации во внешнем пространстве у сверчков Gryllus assimilis // Экспериментальная психология. 2013. Том 6. № 4. С. 79–95.
Полный текст
Изучение специфики самоотражения животных разных таксономических групп является перспективной задачей зоопсихологии и сравнительной психологии. За последние несколько лет авторами настоящей статьи был проведен ряд эмпирических исследований, позволивших выявить особенности самоотражения2 различных позвоночных и беспозвоночных (Филиппова, Хватов, 2011; Хватов, 2010а; 2010б; 2011а; 2011б; Хватов, Харитонов, 2012, 2013). В частности был осуществлен сравнительный анализ особенностей самоотражения тараканов и улиток, как представителей членистоногих и моллюсков соответственно. Показана способность этих животных модифицировать самотражение путем научения. Кроме того, было установлено, что у тараканов отражение внешней среды и самоотражение представлены как слитные в структуре единого образа, т.е. изменение характеристик внешней среды ведет к изменению самоотражения, как и наоборот: модификация характеристик тела насекомого приводит к тому, что оно начинает иначе воспринимать окружающую среду (Хватов, 2011а). Улитки же способны учитывать физические параметры наружной мягкой части своего тела (головы и ноги) при ориентации во внешней среде (Хватов, Харитонов, 2012), а также менять алгоритм движениями этих частей тела при решении задачи на обнаружение обходного пути (Хватов, Харитонов, 2013).
Под самоотражением мы понимаем процесс и результат отражения субъектом своей внутренней объективной реальности: характеристик своего организма – в контексте данного исследования речь будет вестись о размерах и границах собственного тела (Хватов, 2010а).
Настоящее исследование продолжает цикл работ, посвященных изучению особенностей самоотражения беспозвоночных. Объектом исследования были сверчки вида Gryllus assimilis. Такой выбор определялся тем, что морфофизиология и поведение этих насекомых достаточно хорошо изучены.
Целью нашего исследования было выявить, какую сенсорную информацию сверчки учитывают при соотнесении габаритов собственного тела с объектами внешней среды – в частности, при определении возможности проникновения в различные типы отверстий.
Чувствительность сенсорных систем членистоногих чрезвычайно высока (Савельев, 2005; Захваткин, 2012). В основе экстероцептивной чувствительности лежат зрение, механорецепция, хеморецепция, а также слуховая чувствительность (Савельев, 2005). При локомоции для обнаружения препятствий многие членистоногие используют тактильную информацию, для получения которой могут использоваться механорецепторы конечностей (Blaesing, Cruse, 2004; Pick, Strauss, 2005), а также антенн (Camhi, Johnson, 1999; Staudachera et al., 2005).
Насекомые используют антенны для зондирования ближайшего пространства. Сверчки и тараканы обладают длинными антеннами – к примеру, у имаго P. americana они достигают такой же длины, как и все туловище (около 40 мм) и состоят из 120 сегментов (Okada, 2009). На каждой антенне имеется множество сенсилл, доставляющих информацию о тактильных и химических раздражителях (Staudachera et al., 2005).
Антенны используются насекомыми для ориентации собственной локомоции относительно внешних объектов (Camhi, Johnson, 1999; Harley et al., 2009; Okada, Toh, 2000; 2006; Pelletier, McLoed, 1994; Staudacher et al., 2005; Watson et al., 2002; Ye et al., 2003). Когда антенна насекомого соприкасается с неподвижным объектом в пространстве, насекомое может остановиться и затем приблизиться к этому объекту. Данное поведение характерно даже для слепых насекомых (Okada, Toh, 2000; 2006). Этот механизм, связанный с позитивным тигмотаксисом, используется насекомыми при обследовании окружающего пространства – т. н. «движение вдоль стен» (в англоязычной литературе такой элемент поведения иногда обозначается как «wall-following behavior») (Camhi, Johnson, 1999; Wessnitzer et al., 2008). Тараканы Blaberus discoidalis после столкновения с препятствием на пути своего движения при выборе между туннелированием (проникновением под препятствие) или залезанием на препятствие ориентируются на яркость освещения. Однако решающее значение при выборе поведенческого ответа играли сенсорные сигналы, поступавшие от антенн насекомого (Harley et al., 2009).
У сверчков антенны способны различать тактильные текстуры внешних объектов – насекомое будет по-разному вести себя при раздражении антенн биотическими или абиотическими факторами (Okada, Akamine, 2012). Также сверчки тактильно обследуют с помощью антенн движущиеся объекты, первоначально воспринятые визуально (Honegger, 1981; Honegger, Campan, 1989).
С другой стороны, необходимо отметить, что визуальная информация также имеет существенное значение для регуляции поведения сверчков. Они воспринимают поляризованный свет (Brunner, Labhart, 1987) и используют его при ориентации в пространстве в частности, при хоуминге (Beugnon, Campan, 1989; Honegger, Campan, 1989; Weber, 1990; Wessnitzer et al., 2008). Кроме того, эти насекомые способы формировать навыки по пространственной ориентации с опорой на визуальные сигналы (Kieruzel, Chmurzyński, 1982; Beugnon, 1986; Honegger, Campan, 1989; Wessnitzer et al., 2008). Однако эта информация является дополнительной по отношению к тактильной чувствительности при обследовании близко расположенных объектов.
Основываясь на вышеизложенных фактах, мы сформулировали гипотезу настоящего исследования следующим образом: для определения возможности проникновения собственного тела в различные типы отверстий у сверчков решающее значение имеют кинестетические ощущения, поступающие от антенн.
Методика экспериментального исследования
Характеристика испытуемых. В экспериментах использовались 100 сверчков имаго. Животные содержались независимо друг от друга в течение суток в маленьких пластиковых контейнерах при температуре 28 – 30 °C, в качестве корма использовались пшеничные отруби и капуста.
Оборудование. Экспериментальный полигон представлял собой круглую арену диаметром 100 мм. Пол камеры подогревался с помощью термоковрика до температуры 50 °C – аналогичная стимуляция применялась в других исследованиях (Wessnitzer et al., 2008). Такая температура некомфортна для насекомого и стимулирует у него фоботаксис – стремление как можно скорее покинуть зону раздражения (Захваткин, 2012).
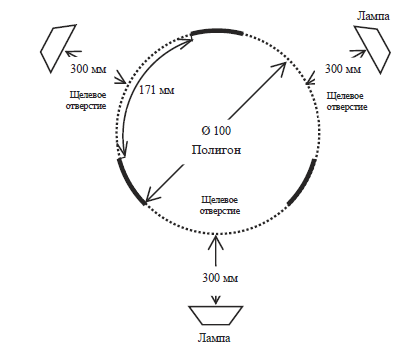
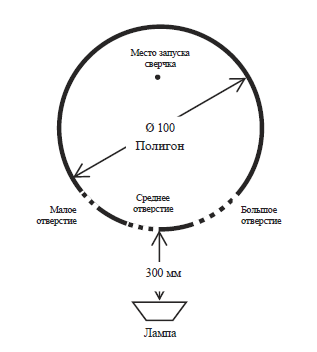
Рис. 2. Схема полигона № 4 – вид сверху
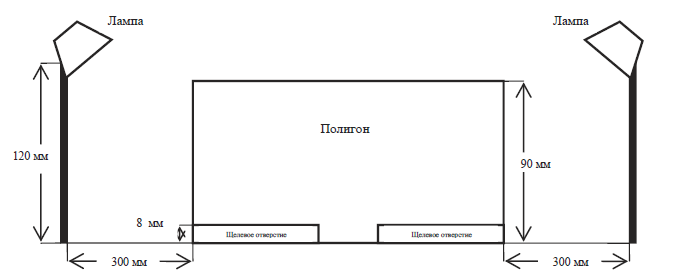
Рис. 3. Схема полигонов № 1, 2, 3 – вид сбоку
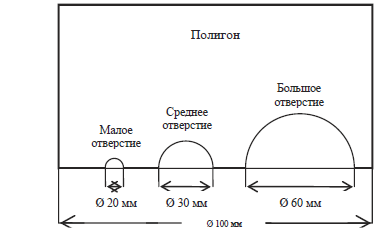
Рис. 4. Схема полигона № 4 – вид ссбоку
Сообразно цели и гипотезе исследования независимой переменной являлся размер отверстия, через которое насекомое могло или не могло покинуть пусковую камеру. Были сконструированы экспериментальные полигоны с различными типами отверстий. Напротив каждого отверстия с внешней стороны на расстоянии 300 мм располагалась люминесцентная лампа, испускающая частично поляризованный свет. Использовалось четыре типа полигонов:
1. Полигон № 1 – в нем между стенами и полом имелось три щелевых отверстия высотой 8 мм и длиной около 171 мм (см. рис. 1 и 3). Сверчки не были способны проникнуть в эти отверстия целиком, но через них легко проникали антенны насекомых.
2. Полигон № 2 – в нем наличествовали аналогичные щелевые отверстия, что и в полигоне № 1, но закрытые металлической сеткой с размером ячеи 0,6 мм. Через отверстия такого размера не способны были проникнуть антенны насекомого, хотя через сетку внутрь пусковой камеры проникал свет.
3. Полигон № 3 – в нем наличествовали аналогичные щелевые отверстия, что и в полигоне № 1, но закрытые металлической сеткой с размером ячеи 2 мм. Через отверстия такого размера антенны сверчка могли проникнуть в том случае, если движение ими совершалось под прямым углом по направлению к сетке, в случае если же движение шло по касательной, то антенны могли отразиться от сетки. Иначе говоря, проникновение антенны через отверстие было вероятностным.
4. Полигон № 4 – в нем имелись три полукруглых отверстия различного диаметра:
20 мм (далее – маленькое отверстие), 30 мм (далее – среднее отверстие) и 60 мм (далее – большое отверстие), расположенные на одном конце полигона (см. рис. 2 и 4). Тело сверчка было способно проникнуть в большие и средние отверстия, через маленькое отверстие проникали только его антенны. В этом полигоне использовалась одна лампа, установленная с внешней стороны напротив трех отверстий.
Процедура эксперимента. Эксперимент состоял из четырех серий с применением одного из четырех полигонов. В экспериментальной серии каждый из 100 сверчков запускался внутрь полигона 1 раз. Соответственно, каждое насекомое тестировалось всеми четырьмя полигонами по одному разу. Сверчок запускался внутрь полигона в случайном месте и проводил там 2 минуты. Фиксировалась траектория передвижения испытуемого внутри полигона, количество попыток проникновения в отверстия, а также то, совершаются ли эти попытки только после того, как в соответствующее отверстие проникнут антенны насекомого, или же они совершаются даже в том случае, если антенны предварительно не проникли через искомое отверстие.
Эмпирическим критерием, подтверждающим выдвинутую гипотезу, должен был являться тот факт, что сверчки достоверно чаще совершают попытки проникновения в отверстия лишь после того, как в эти отверстия уже проникнут их антенны.
Несколько иначе организовывалась экспериментальная серия с применением полигона № 4. Сверчок запускался в него в определенном месте головой в сторону отверстий (см. Рис. 4). Каждая экспериментальная проба длилась до тех пор, пока насекомое не покидало экспериментальный полигон через среднее или большое отверстие. Фиксировалась траектория движения испытуемого, количество выходов через средние и большие отверстия, а также количество попыток проникновения в малое отверстие. Решалась задача выявить, в какие из трех типов отверстий сверчки предпочитают проникать. Это позволяло определить роль зрительной системы сверчков в решении экспериментальной задачи. В случае достоверно большего числа проникновений в большие и средние отверстия в сравнении с попытками проникновения в маленькие при том, что насекомые предварительно не обследовали данные отверстия с помощью антенн, можно утверждать, что в данном поведении они опираются именно на визуальную информацию.
Эксперимент проводился в течение 10 дней. Каждый день тестировалось 10 сверчков с помощью всех полигонов: насекомое последовательно помещали в каждый из них. Интервал между пробами составлял две минуты. После каждой пробы полигоны тщательно очищались, чтобы исключить влияние различных химических агентов на поведение сверчков.
Характеристика фиксируемых элементов поведения.
Под «проникновением антенн в отверстие» имелась в виду ситуация, когда обе антенны насекомого проникали через то или иное отверстие и выходили за пределы пусковой камеры более чем на 2 мм.
Под «попыткой проникновения в отверстия» имелась в виду ситуация, когда сверчок, прижавшись головой или же другими сегментами тела к стенке или сетке, окаймляющей данное отверстие, совершал толкательные и/или роющие движения ногами в сторону отверстия более 1 сек. подряд. Также фиксировалось длительной каждой попытки проникновения.
Под «выходом из экспериментального полигона» имелась в виду ситуация, когда тело сверчка целиком покидало полигон № 4 через среднее или большое отверстие.
Кроме того, фиксировались такая форма поведения как «попытка залезть на стену», под которой понималась ситуация, когда сверчки ставили свое тело вертикально, опираясь на стену передними парами ног, активно перебирая ими, поднимали голову вверх и совершали толкательные движения задней парой ног. Таки попытки были неэффективны, поскольку поверхность стен была гладкой и насекомые не могли забраться на них.
В полигонах, где применялась сетка (полигоны № 2 и № 3), фиксировалось количество тактильных «контактов с сеткой»: ситуаций, когда обе антенны соприкасались с сеткой, но не проникали за ее пределы, хотя такое проникновение было возможно.
В ряде случаев после контакта с сеткой или после проникновения антенн в отверстие насекомые не совершали каких-либо из вышеуказанных двигательных операций (попытки проникновения в отверстие или попытки залезть на стену), а продолжали свое движение вдоль стены или же снова уходили к центру полигона. Такие поведенческие проявления также фиксировались и относились к категории «уход в сторону».
Помимо подсчета общего количество описанных элементов поведения строились этограммы.
Анализ данных. Поведение сверчка внутри полигона в каждой пробе фиксировалось с помощью камеры (Sony HandyCam HDR-CX220), закрепленной над полигоном на высоте 30 см. Полученные количественные данные обрабатывались с помощью методов непараметрической статистики с применением программы STATISTICA 8,0.
Результаты экспериментального исследования
Экспериментальная серия № 1 (полигон № 1). После помещения в полигон сверчки сразу же начинали движение от центра к краям. Осуществляя поисковые движения антеннами, ощупывая ими стены, насекомые проникали антеннами в щелевые отверстия. Среднее количество проникновений антенн в щелевые отверстия у одного испытуемого за одну пробу составляло 7,97 (SD=3,14), среднее количество следовавших за этим попыток проникновения сверчка в данное отверстие составляло 7,04 (SD=2,02), т.е. в 88% случаев после проникновения собственных антенн в щелевые отверстия сверчки пытались проникнуть в них и всем телом. Уходы в сторону и попытки залезть на стену после проникновения антенн в отверстие встречались гораздо реже: m=0,75 (SD=1,27) и m=0,18 (SD=0,57), соответственно. Средняя продолжительность попытки проникновения в отверстие составлялат 6,71 с (SD=2,90).
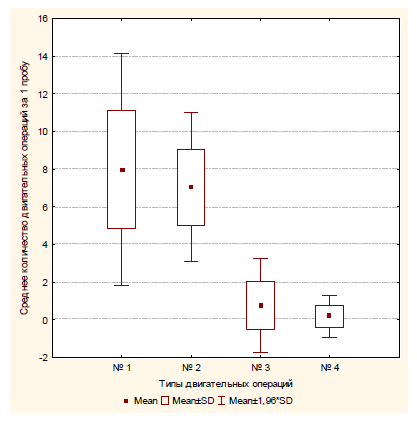
Рис. 5. Среднее количество различных операций, совершаемое испытуемым за одну пробу в первой экспериментальной серии: № 1 – проникновение антенн в отверстие; № 2 – попытка проникновения в отверстие; № 3 – уход в сторону; № 4 – попытка залезть на стену
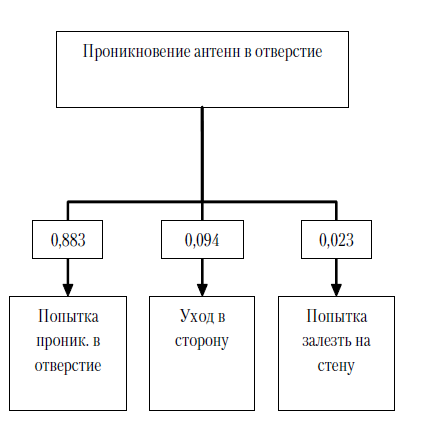
Рис. 6. Этограмма сверчков для первой экспериментальной серии
После запуска внутрь полигона сверчки сразу же направлялись к стене и около 95% времени проводили, двигаясь вдоль стен полигона и обследуя их антеннами. В среднем около 47 с (из общих 120 секунд, проводимых внутри полигона за одну пробу) сверчки тратили на попытки проникновения в щелевые отверстия. Попытка проникновения представляла собой следующее: после проникновения антенны в щелевое отверстие сверчок приближался к нему вплотную головой, затем прижимал голову, переднеи среднегрудь к полу, благодаря чему в щелевое отверстие проникали уже не только антенны, но и членики ротового аппарата (в частности, пальпы), совершая при этом толкательные движения первыми двумя парами ног (в некоторых случаях такие толчки осуществлялись и с помощью задней прыгательной пары ног).
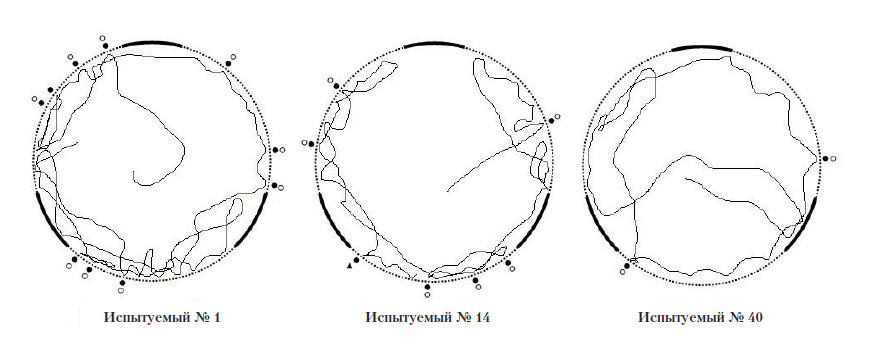
Рис. 7. Примеры траектории движения испытуемых в полигоне № 1.
Условные обозначения: черный кружок – проникновение антенн в отверстие, светлый кружок – попытка проникновения в отверстие, черный треугольник – попытка залезть на стену
В полигоне № 1 после проникновения обеими антеннами в щелевое отверстие сверчки достоверно чаще совершали попытки проникнуть в это отверстие всем телом, нежели какие-либо другие действия («уход в сторону» и «попытка залезть на стену»): χ2=1090,95 (df=2, p<0,01).
Экспериментальная серия № 2 (полигон № 2). После помещения в полигон сверчки вели себя аналогичным образом, что и в первой экспериментальной серии. Насекомые ощупывали антеннами стены и сетку, закрывающую щелевые отверстия. Среднее количество контактов антенн с сеткой 11,24 (SD=3,21), проникновения антенн в отверстия сетки практически отсутствовали (m=0,02, SD=0,14), в большинстве случаев после контакта с сеткой сверчки либо уходили в сторону (m=5,55, SD=2,96), либо совершали попытки залезть на стену (m=5,66, SD=2,07). Попытки проникновения сверчков в щелевые отверстия после контакта антенн с сеткой также практически отсутствовали (m=0,03, SD=0,07) – менее 0,01 % от всех действий, совершаемых после контакта с сеткой.
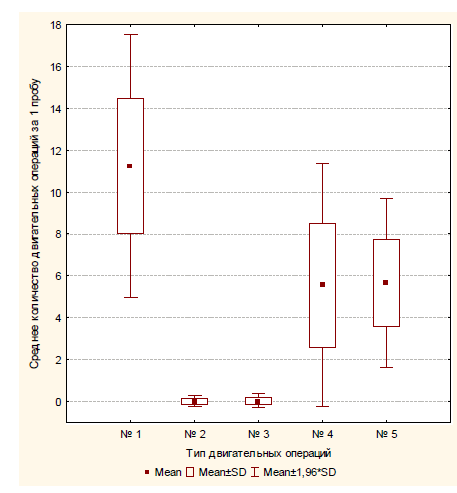
Рис. 8. Среднее количество различных операций, совершаемое испытуемым за одну пробу во второй экспериментальной серии: № 1 – контакт с сеткой; № 2 – проникновение антенн в отверстие; № 3 – попытка проникновения в отверстие; № 4 – уход в сторону; № 5 – попытка залезть на стену
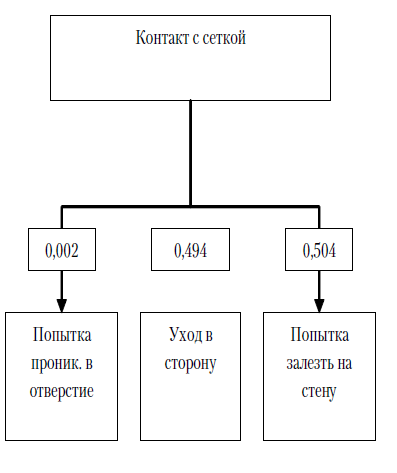
Рис. 9. Этограмма сверчков для второй экспериментальной серии
Так же как и в экспериментальной серии № 1, большую часть времени сверчки провели в процессе обследования антеннами стен по периметру полигона.
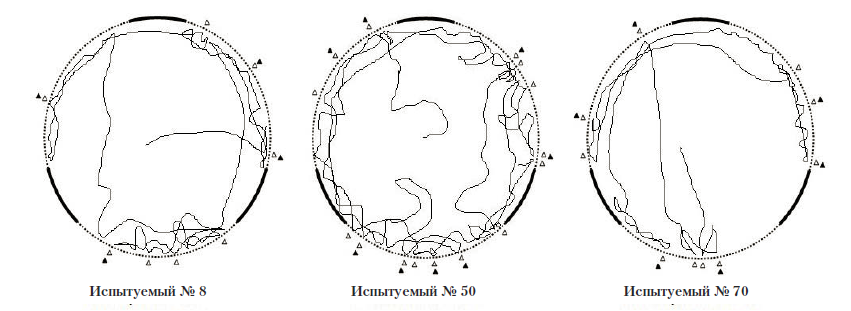
Рис. 10. Примеры траектории движения испытуемых в полигоне № 2.
Условные обозначения: светлый треугольник – контакт антенн с сеткой; черный треугольник – попытка залезть на стену
В полигоне № 2 после контакта обеими антеннами с сеткой (антенны практически не проникали в ячейки сетки) сверчки достоверно чаще совершали такие действия, как «уход в сторону» или «попытка залезть на стену», нежели пытались проникнуть в закрытое сеткой щелевое отверстие: χ2=553,20 (df=2, p<0,01).
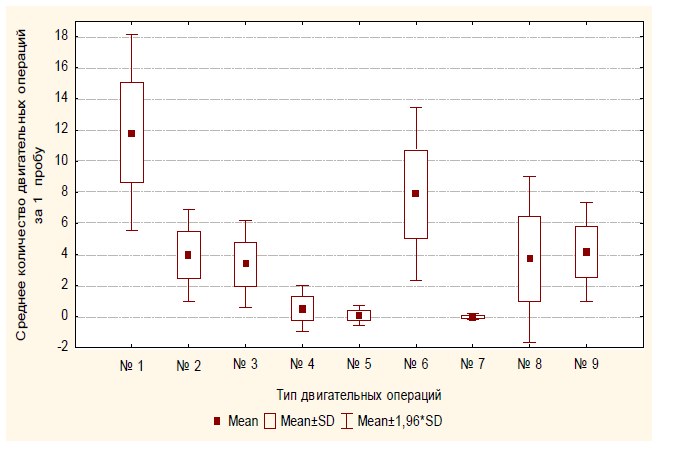
Рис. 11. Среднее количество различных операций, совершаемое испытуемым за одну пробу в третьей экспериментальной серии: № 1 – контакт с сеткой; № 2 – проникновение антенн в отверстие; № 3 – № 5 – действия, совершенные после проникновения антенн в отверстие: № 3 – попытка проникновения в отверстие; № 4 – уход в сторону; № 5 – попытка залезть на стену; № 6 – ситуации, когда антенны не проникали в отверстие после их контакта с сеткой; № 7 – № 9 – действия, совершенные в ситуациях, когда при соприкосновении с сеткой антенны не проникали в отверстие: № 7 – попытка проникновения в отверстие; № 8 – уход в сторону; № 9 – попытка залезть на стену
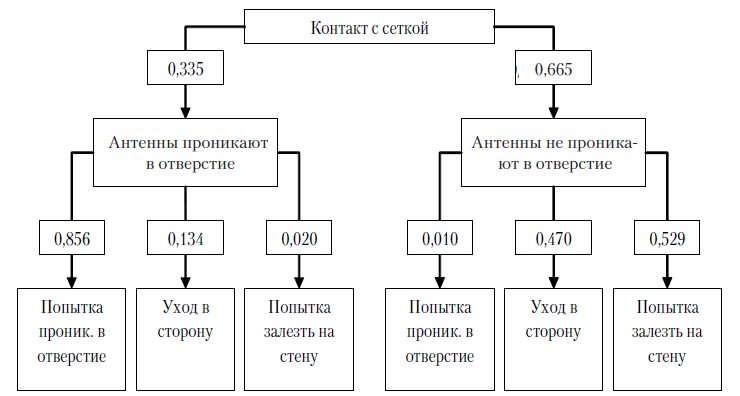
Рис. 12. Этограмма сверчков для третьей экспериментальной серии
Затем с помощью T-критерия Вилкоксона были сопоставлены между собой эмпирические распределения количества попыток проникновения в щелевые отверстия, продемонстрированные испытуемыми в ходе экспериментальных серий № 1 и № 2. Эмпирические распределения достоверно отличались друг от друга (p<0,01). Таким образом, в ходе первой экспериментальной серии сверчки достоверно чаще совершали попытки проникновения в щелевые отверстия.
Экспериментальная серия № 3 (полигон № 3). В ходе данной экспериментальной серии в процессе обследования сетки, закрывающей щелевое отверстие (среднее количество контактов с сеткой за 1 пробу – 11,8; SD=3,21), в 33,5% случаев сверчки проникали антеннами в ячейки сетки за пределы щелевого отверстия (среднее количество проникновений антенн в отверстие за 1 пробу – 4,0; SD=1,5), в 66,5% случаев антенны не проникали за пределы щелевого отверстия. После проникновения антенны в щелевое отверстие в 85,6% случаев сверчки совершали попытки проникнуть в данное отверстие всем телом (среднее количество попыток проникновения в отверстие – 3,4; SD=1,4); другие действия совершались значительно реже: уход в сторону в 13,4% случаев (среднее за одну пробу – 0,5; SD=0,76), попытка залезть на стену в 2% (среднее за одну пробу – 0,08; SD=0,34). Иначе разворачивалось поведение сверчков в ситуациях, когда после контакта антеннами с сеткой сверчки не проникали ими в щелевое отверстие (это происходило в 66,5% случаев, в среднем 7,88 раз за одну пробу, SD=2,84): только в 0,1% таких случаев сверчки совершали попытки проникновения в щелевые отверстия, гораздо чаще они уходили в строну (в 47% случаев, в среднем
3,7 раз за одну пробу, SD=2,72) или совершали попытку залезть на стену (в 52,9% случаев, в среднем 4,17 раз за одну пробу, SD=1,63). Средняя продолжительность попытки проникновения в отверстие составляла 6,65 с (SD=2,62).
Необходимо отметить, что в полигоне № 3 попытки проникновения в щелевые отверстия имели некоторые качественные отличия от тех попыток, что наблюдались у сверчков в полигоне № 1. В третьей экспериментальной серии после проникновения антенн в щелевое отверстие через ячейки сетки при последующем приближении к ней в 90% случаев сверчки дотрагивались до сетки пальпами и ощупывали ее ими (в отличие от попыток проникновения в первой экспериментальной серии, когда пальпы также проникали в отверстие). Затем сверчок совершал роющие движения передней парой конечностей по поверхности пола, прилегающего к сетке, а также по поверхности самой сетки; второй и третьей парой конечностей он совершал толкательные движения, аналогичные тем, что наблюдались в первой экспериментальной серии. При этом пальпами насекомое совершало расталкивающие движения в ячейках между проволокой сетки, иногда кусая проволоку мандибулами.
В полигоне № 3 после проникновения обеими антеннами в щелевое отверстие сверчки достоверно чаще совершали попытки проникнуть в это отверстие всем телом, нежели какие-либо другие действия («уход в сторону» и «попытка залезть на стену»): χ2=483,45 (df=2, p<0,01). Также в случае не проникновения антенн в отверстия сверчки достоверно чаще совершали такие действия, как «уход в сторону» или «попытка залезть на стену», нежели пытались проникнуть в щелевое отверстие: χ2=395,21 (df=2, p<0,01).
Отдельно следует отметить, что испытуемые достоверно чаще совершали попытки проникновения в щелевое отверстие всем телом в тех случаях, когда в эти отверстия предварительно проникали их антенны, нежели в случаях, когда этого не происходило: χ2=334,03 (df=2, p<0,01).
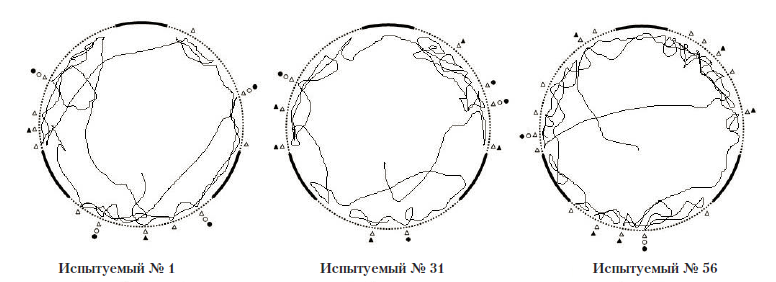
Рис. 13. Примеры траектории движения испытуемых в полигоне № 3.
Условные обозначения: черный круг – проникновение антенн в отверстие, светлый круг – попытка проникновения в отверстие светлый треугольник – контакт антенн с сеткой; черный треугольник – попытка залезть на стену
Экспериментальная серия № 4 (полигон № 4). После помещения в полигон сверчки сразу направлялись в сторону одного из отверстий и покидали полигон в среднем за 11,6 с (SD=2,7), т.е. проводили внутри значительно меньше времени, нежели в первых трех экспериментальных сериях. Общее количество попыток проникновения в маленькие отверстии за всю серию – 22 (m=0,22; SD=0,52), общее количество проникновений в средние отверстия – 37 (m=0,37; SD=0,48), общее количество проникновений в большие отверстия – 63 (m=0,63; SD=0,48). Следует отметить, что все попытки проникновения в маленькие отверстия совершались после предварительного проникновения в них антенн испытуемых. Было зафиксировано три случая, когда при обследовании малых отверстий сверчки не проникали в них антеннами и сразу после этого двигались в сторону других отверстий (см. рис. 15 – траектория движения испытуемого № 39). Средняя продолжительность попытки проникновения в малое отверстие составляла 3,77 с (SD=1,27).
Сверчки достоверно чаще проникали в большое отверстие, нежели в среднее или совершали попытку проникнуть в малое: χ2=21,16 (df=2, p<0,01).
Анализ результатов исследования
Данные, полученные в ходе первых трех экспериментальных серий, свидетельствуют, что при определении проницаемости или непроницаемости отверстия для их тела сверчки опираются на информацию, поступающую от их антенн – эта информация является определяющей для решения данной задачи. В случае проникновения антенн в отверстия сверчки достоверно чаще пытались проникнуть туда всем телом, нежели совершать какие-либо другие двигательные операции, что объясняется их мотивацией покинуть пусковую камеру, условия которой некомфортны для этих животных. Данная закономерность сохраняется и в том случае, если отверстия, через которые проникают антенны, являются настолько маленькими (в третьем полигоне размер ячеи сетки составлял 2 мм), что обе антенны проникают в разные ячейки. Однако субъективно для насекомого это, очевидно, означает, что перед ним находится открытое пространство, в котором может продвигаться все его тело. При этом, очевидно, насекомое игнорирует кинестетические сигналы, поступающие от рецепторов других частей тела – в частности тех мест, что соприкасаются с краями отверстия и препятствуют продвижению вперед.
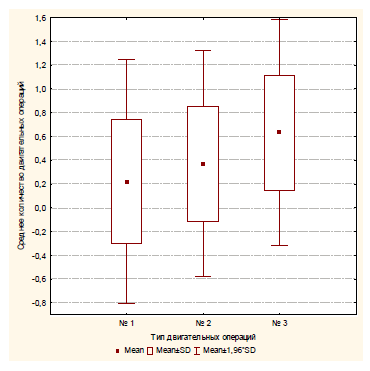
Рис. 14. Среднее количество различных операций, совершаемое испытуемым за одну пробу в четвертой экспериментальной серии: № 1 – попытка проникновения в малое отверстие; № 2 – проникновение в среднее отверстие; № 3 – проникновение в большое отверстие
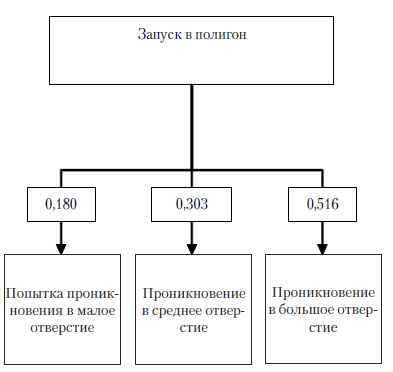
Рис. 15. Этограмма сверчков для четвертой экспериментальной серии
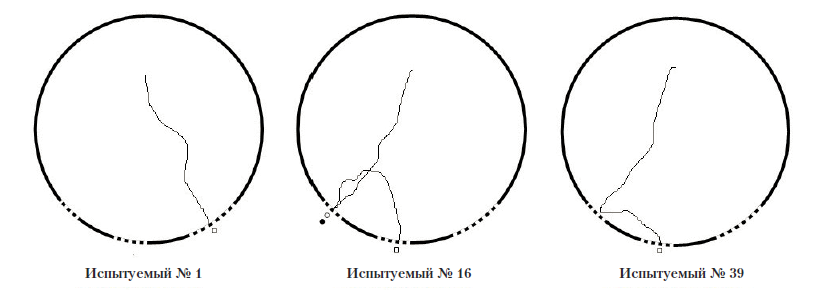
Рис. 16. Примеры траектории движения испытуемых в полигоне № 4.
Условные обозначения: черный круг – проникновение антенн в отверстие, светлый круг – попытка проникновения в отверстие, светлый квадрат – выход из полигона
Необходимо выделить два различных способа проникновения в отверстия, продемонстрированных сверчками. Первый способ наблюдался при поведении в полигоне № 1 (см. описание экспериментальной серии № 1) – условно его можно назвать «попыткой пролезть в отверстие», не сопровождающейся роющими движениями. Второй способ наблюдался при поведении в полигоне № 3 (см. описание экспериментальной серии № 3) – он представлял собой попытку прорыть себе путь, расширив отверстие, что, вероятно, обусловлено тем, что даже при проникновении антенн через ячейки сетки, сверчки все же ощущали ее с помощью пальп. Специфика второго способа проникновения в отверстия объясняется экологией сверчков: большинство видов этих насекомых являются наземными жителями и часто роют норы в грунте (Grantzy, Hustert, 1989). Так, в частности, при откладке яиц, оказываясь на влажном грунте, самки разрывают грунт передними парами конечностей, также перебирая песчинки пальпами. Иными словами, в природных условиях для сверчков не типична ситуация, когда то или иное отверстие является непроницаемым для их тела – в большинстве случаев его можно расширить с помощью роющих движений. Однако в обоих случаях (и при первом, и при втором способе проникновения в отверстия), туда предварительно проникали антенны насекомых. В случаях, когда антенны не проникали за пределы сетки (в полигонах № 2 и № 3), т.е. натыкались на нее как на препятствие и затем отклонялись в своем движении вверх, вниз или вбок, сверчки не совершали роющих движений, но зачастую напротив – пытались забраться на стену.
В четвертой экспериментальной серии сверчки достоверно чаще проникали в большие отверстия, нежели пытались проникнуть в малые. Однако этот факт не свидетельствует о том, что при определении возможности проникновения собственного тела в отверстия полигона № 4 сверчки опирались именно на визуальную информацию. При определении траектории своего движения, ориентируясь на свет (Beugnon Campan, 1989; Honegger, Campan, 1989; Weber, 1990; Wessnitzer et al., 2008), они, вероятно, чаще двигались в направлении большего отверстия, поскольку оно пропускает больший поток света. Тем не менее, в случаях приближения к малому отверстию, обследования его антеннами и при условии, проникновения антенн в это отверстие, сверчки совершали попытки проникновения в них всем телом (было зафиксировано 22 таких случая). С другой стороны, когда антенны не проникали в малые отверстия при их обследовании, сверчки не совершали попыток проникнуть в них всем телом (было зафиксировано 3 таких случая). Таким образом, визуальная информация задавала только общую ориентацию движений сверчков в направлении к свету, но определение возможности или невозможности проникновения в те или иные отверстия осуществлялось на основе кинестетических сигналов, поступающих от антенн.
Соответственно, полученные нами данные подтверждают выдвинутую гипотезу исследования. Сверчки используют антенны в качестве зондов, доставляющих информацию о наличии или отсутствии препятствий на пути следования насекомого. В частности, эта информация играет определяющую роль при принятии решения о возможности или невозможности проникновения собственного тела в те или иные отверстия.
Невзирая на то, что, в соответствии с периодизацией развития психики А. Н. Леонтьева (1972) и более поздними ее модификациями (см., например: Филиппова, 2012), насекомые относятся к перцептивной стадии эволюции психики (т. е. они способны строить интермодальные психические образы на основе интеграции информации, поступающей от разных сенсорных систем), у сверчков самоотражение, как и отражение внешнего мира фактически остается мономодальным в том смысле, что выбор определенной поведенческой программы определяется наиболее сильным раздражением, поступающим от определенной части тела (Савельев, 2005). У насекомых ориентация поведения относительно внешних объектов регулируется центральным комплексом (англоязычный термин «the central complex»), расположенным в протоцеребруме, где осуществляется обработка тактильной, химической и визуальной информации (Vitzthum et al., 2002; Ritzmann et al., 2008; Heinze, Homberg, 2007; Homberg, 2004). В ходе нашего эксперимента насекомые ориентировались на тактильные и визуальные сигналы, однако, очевидно, в выборе поведения конкретной ситуации решающую роль играют именно тактильные сигналы, поступающие от антенн. Эти данные согласуются с данными, ранее полученными на тараканах (Harley et al., 2009).
Опираясь на концепцию самоотражения животных и человека, результаты настоящего экспериментального исследования мы можем объяснить следующим образом. Сверчки, как и другие представители насекомых – тараканы (Хватов, 2011a), лишены схемы собственного тела в качестве целостной самостоятельной когнитивной структуры. Такая схема складывается у них ситуативно в качестве элемента определенной перцептивной системы (целостного образа ситуации), формирующегося в процессе взаимодействия субъекта с конкретными условиями внешней среды. В ситуации нашего эксперимента сверчки принимали решение о возможности или невозможности проникновения собственного тела в отверстие, т.е. формировали структуру образа ситуации со слитыми внутри нее самоотражением и отражением внешнего мира, в процессе обследования антеннами данного отверстия. Если целостный образ ситуации и являлся перцептивным (в частности, в его структуре интегрируется визуальная и кинестетическая информация) – самоотражение как компонент образа ситуации остается сенсорным, поскольку формируется на основе кинестестической информации, поступающей от антенн и пальп, в то время как сигналы от других частей тела – в частности тех, что соприкасаются с краями отверстия – игнорируются. Фактически сверчок субъективно не соотносит физические параметры собственного тела с размером отверстия, определяющим фактором является то, проникают или не проникают в него антенны, независимо от того, насколько малым оказывается это отверстие. Этим поведение сверчков Gryllus assimilis отличается от поведения улиток Achatina fulica, способных соотносить физические параметры наружной мягкой части своего тела (головы и ноги) с характеристиками внешних объектов при ориентации в окружающей среде (Хватов, Харитонов, 2012).
Выводы
На основании проведенного анализа результатов настоящего исследования мы можем констатировать следующее:
-
Для определения возможности проникновения собственного тела в различные типы отверстий у сверчков решающее значение имеют кинестетические ощущения, поступающие от антенн;
-
В зависимости от совокупности двигательных операций попытки проникновения в отверстия у сверчков можно подразделить на два типа: пролезание через отверстие и протискивание через отверстие;
-
Способ проникновения в отверстия определяется кинестетической информацией, поступающей от ротовых придатков – в первую очередь пальп.
-
Эти выводы нуждаются в дополнительной экспериментальной проверке. В частности, перспективным представляется эксперимент с поиском выхода из экспериментальной установки при варьировании как размерами отверстий, так и размерами тела сверчков – по аналогии с тем экспериментом, что ранее был организован нами на тараканах (Хватов, 2011a). Также возможна постановка эксперимента с варьированием в качестве независимой переменной направлением и яркостью света, вплоть до полного отсутствия освещения.
Литература
- Захваткин Ю. А. Курс общей энтомологии: Учебник. Изд. 3-е. М.: ЛИБРОКОМ, 2012.
- Леонтьев А. Н. Проблемы развития психики. 3-е изд. М.: Моск. гос. университет, 1972.
- Савельев С. В. Происхождение мозга. М.: ВЕДИ, 2005.
- Хватов И. А. Особенности самоотражения у животных на разных стадиях филогенеза. Дисс. канд. психол. наук. М., 2010а.
- Хватов И. А. Специфика самоотражения у вида Periplaneta americana // Экспериментальная психология. 2011а. № 1. C. 28–40.
- Хватов И. А. Эмпирическое исследование восприятия самих себя у понгид (на примере рисования перед зеркалом у орангутанов) // 125 лет Московскому психологическому обществу: Юбилейный сборник РПО: В 4-х томах / Отв. ред. Богоявленская Д. Б., Зинченко Ю. П. М.: МАКС Пресс, 2011б. Т. 2. С. 46–47.
- Хватов И. А. Эмпирическое исследование проблемы филогенетических предпосылок становления самосознания // Знание. Понимание. Умение. 2010б. № 2. С. 242–247.
- Хватов И. А., Харитонов А. Н. Модификация плана развертки собственного тела в процессе научения при решении задачи на нахождение обходного пути у улиток вида Achatina fulica // Экспериментальная психология. 2013. № 2. С. 101–114.
- Хватов И. А., Харитонов А. Н. Специфика самоотражения у вида Achatina fulica // Экспериментальная психология. 2012. № 3. C. 96–107.
- Филиппова Г. Г. Зоопсихология и сравнительная психология: учеб. пособие для студентов вузов. 6-е изд., перераб. М.: Академия, 2012.
- Филиппова Г. Г., Хватов И. А. Специфика экспериментального метода в зоопсихологии на примере исследования самоотражения у животных на интеллектуальной стадии развития психики // Современная экспериментальная психология: В 2 т. / Под ред. В. А. Барабанщикова. М.: Изд-во «Институт психологии РАН», 2011. Т. 1. С. 499–511.
- Beugnon G. Learned orientation in landward swimming in the cricket Pteronemobius lineolatus // Behav. Process. 1986. № 12. P. 215–226. doi: 10.1016/0376-6357(86)90037-9.
- Beugnon G., Campan R. Homing in the field cricket Gryllus campestris // J. Insect Behav. 1989. № 2. P. 187–198. doi: 10.1007/BF01053291
- Blaesing B., Cruse H. Stick insect locomotion in a complex environment: climbing over large gaps // J. Exp. Biol. 2004. № 207. P. 1273–1286.
- Brunner D., Labhart T. Behavioural evidence for polarization vision in crickets // Physiol. Entomol. 1987. № 12. P. 1–10.
- Camhi J. M., Johnson E. N. High-frequency steering maneuvers mediated by tactile cues: Antennal wallfollowing in the cockroach // Journal of Experimental Biology A. 1999. № 202. P. 631–643.
- Grantzy W., Hustert R. Mechanoreceptors in behavior // Cricket Behavior and Neurobiology / Eds. F. Huber, T. Edwin Moore, W. Loher. Ithaca. Cornell University Press: 1989. P. 198–226.
- Heinze S., Homberg U. Maplike representation of celestial E-vector orientations in the brain of an insect // Science. 2007. № 315. P. 995–997.
- Harley C. M., English B. A., Ritzmann R. E. Characterization of obstacle negotiation behaviors in the cockroach, Blaberus discoidalis // J. Exp Biol. 2009. № 212 (10). P. 1463–1476.
- Homberg U. Multisensory processing in the insect brain // Methods in Insect Sensory Neuroscience / Ed. T. A. Christensen. CRC Press, Boca Raton, London, New York, Washington, DC, 2004. P. 3–25.
- Honegger H.-W. A preliminary note on a new optomotor response in crickets: Antennal tracking of moving targets // Journal of comparative physiology. 1981. V. 142. № 3. P. 419–421.
- Honegger H.-W., Campan R. Vision and Visually Guided Behavior // Cricket Behavior and Neurobiology / Eds. F. Huber, T. Edwin Moore, W. Loher. Ithaca: Cornell University Press, 1989. P. 147–177.
- Kieruzel M., Chmurzyński J. A. Visual preferences for certain flat patterns in the house cricket and their conditionally acquired changes // Biol. Behav. 1982. № 7. P. 119–135.
- Nishino H., Nishikawa M., Yokohari F., Mizunami M. Dual, multilayered somatosensory maps formed by antennal tactile and contact chemosensory afferents in an insect brain // Journal of Comparative Neurology, 2005. № 493(2). P. 291–308.
- Okada J. Cockroach antennae // Scholarpedia. 2009. № 4(10). P. 6842. doi: 10.4249/scholarpedia.6842. Okada J., Akamine S. Behavioral response to antennal tactile stimulation in the field cricket Gryllus bimaculatus // Journal of Comparative Physiology A. 2012. V. 198. № 7. P. 557–565.
- Okada J., Toh Y. Active tactile sensing for localization of objects by the cockroach antenna // Journal of Comparative Physiology A. 2006. V. 192. № 7. P. 715–726.
- Okada J., Toh Y. Peripheral representation of antennal orientation by the scapal hair plate of the cockroach Periplaneta americana // J. Exp. Biol. 2001. № 204. P. 4301–4309.
- Okada J., Toh, Y. The role of antennal hair plates in object-guided tactile orientation of the cockroach (Periplaneta americana) // Journal of Comparative Physiology. 2000. № 186. P. 849–857.
- Pelletier Y., McLoed C. Obstacle perception by insect antennae during terrestrial locomotion // Physiol. Entomol. 1994. V. 19. № 4. P. 360–362.
- Pick S., Strauss R. Goal-driven behavioral adaptations in gap-climbing Drosophila // Current Biology. 2005. V. 15. № 16. P. 1473–1478.
- Ritzmann R. E., Ridgel, A. L. and Pollack A. J. Multi-unit recording of antennal mechano-sensitive units in the central complex of the cockroach Blaberus discoidalis // J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2008. № 194. P. 341–360.
- Ruppert E. E., Fox R. S., Barnes R. D. Invertebrate Zoology; a Functunal Evolutionary Approach. Stamford: Cengage Learning, 2009.
- Staudacher E. M., Gebhardtb M., Dürrc V. Antennal Movements and Mechanoreception: Neurobiology of Active Tactile Sensors // Advances in Insect Physiology. 2005. V. 32. P. 49–205.
- Vitzthum H., Muller M., Homberg U. Neurons of the central complex of the locust Schistocerca gregaria are sensitive to polarized light // J. Neurosci. 2002. № 22(3). P. 1114–1125.
- Watson J. T., Ritzmann R. E., Zill S. N., Pollack A. J. Control of obstacle climbing in the cockroach, Blaberus discoidalis. I. Kinematics // Journal of Comparative Physiology A. 2002. V. 188. № 1. P. 39–53.
- Weber T. Phonotaxis and visual orientation in Gryllus campestris: behavioural experiments // Sensory systems and communication in arthropods : including the first comprehensive collection of contributions by Soviet scientists / Ed. F. G. Gribakin. Boston: Birkhauser Verlag, 1990. P. 377–386.
- Wessnitzer J., Mangan M., Webb B. Place memory in crickets // Proc. R. Soc. B. 2008. V. 275. № 1637. P. 915–921. doi: 10.1098/rspb.2007.1647
- Ye S., Leung V., Khan A., Baba Y., Comer C. M. The antennal system and cockroach evasive behavior. I. Roles for visual and mechanosensory cues in the response // Journal of Comparative Physiology A. 2003. V. 189. № 2. P. 89–96.
Информация об авторах
Метрики
Просмотров
Всего: 2980
В прошлом месяце: 11
В текущем месяце: 26
Скачиваний
Всего: 673
В прошлом месяце: 1
В текущем месяце: 1