![]()
ХВАТОВ И. А. , Московский институт психоанализа, Москва, Россия, e-mail: ittkrot1@gmail.com
СОКОЛОВ А. Ю.[*] [†], АНО Лаборатория-студия «Живая Земля», Москва, Россия, e-mail:apophisking@mail.ru
ХАРИТОНОВ А. Н.[‡], Институт психологии РАН; Московский государственный психологопедагогический университет, Москва, Россия, e-mail: ankhome47@list.ru
Схема собственного тела у гигантских синеязыких сцинков Tiliqua gigas изучалась с помощью методики, в которой животным было необходимо учитывать увеличенные границы собственного тела при нахождении пути движения через отверстия различных диаметров в экспериментальной установке. Было установлено, что сцинки способны научиться учитывать естественные границы собственного тела при взаимодействии с объектами окружающей среды. Полученные результаты свидетельствуют также, что при увеличении границ тела сцинки способны модифицировать схему тела - учитывать объективные изменения габаритов собственного тела при ориентации в окружающей среде. В эксперименте это выражалось в снижении количества неуспешных попыток проникновения, совершенных сцинками с увеличенными границами их головы, в отверстия, через которые ранее их неизмененное тело могло проникнуть. При изменении расположения отверстий в экспериментальной установке сцинки оказались способны переносить ранее сформированный навык в новую ситуацию.
Введение
Схема тела - это совокупность двигательных навыков и способностей, позволяющих осуществлять различные движения, а также представление о физических характеристиках собственного тела (его границах, весе, взаиморасположении отдельных членов) (Head, Holmes, 1911; Gallagher, Cole, 1995). Схема тела предполагает соотнесение субъектом характеристик собственного тела с характеристиками окружающих объектов и необходима ему для пространственной ориентации и планирования движений (Head, Holmes, 1911; Graziano et al., 1994; Gallagher, Cole, 1995), т. е. для «принятия себя в расчет» (Столин, 1983). В отличие от образа тела, схема тела является неосознаваемым феноменом (Gallagher, Cole, 1995). Существует точка зрения, что схема тела является филогенетически наиболее ранней ступенью развития всех прочих представлений о себе, включая самосознание (Столин, 1983). Значительная часть исследований схемы тела и возможностей ее модификации проводится на людях, других приматах и некоторых других видах млекопитающих (Herman et al., 2001; Maravita, Iriki, 2004; Johnson-Frey, 2004). Недавние исследование показали, что схема тела обладает высокой пластичностью и способна интегрировать в свою структуру внешние объекты, находящиеся в физическом контакте с индивидом, например, орудия, используемые им (Carlson et al., 2010; Gozli, Brown, 2011; Ritchie, Carlson, 2013; Costantini et al., 2014; Moeller et al., 2015; Garbarinia et al., 2015). Эти факты согласуются с идеей рассмотрения орудия как зонда (Леонтьев, 1975; Тхостов, 2002). Сообразно изменению схемы тела происходит и модификация субъективного восприятия окружающей реальности - эгоцентрической системы координат (Барабанщиков, 2002; Berti, Frassinetti, 2000; Costantini et al., 2014; Giglia et al., 2015).
Между тем, в научной литературе отсутствуют публикации, описывающие исследования феномена схемы тела и ее изменений у большинства видов позвоночных. Большинство поведенческих аспектов, связанных с феноменом схемы тела, не рассматриваются в их связи с феноменами самосознания и/или Я-концепции, как с онтологической, так и с эволюционной точек зрения. Исключением является крупное направление исследований способности животных к самоузнаванию в зеркале (англ. «Mirror test»), развивающееся начиная с классических экспериментов Г. Гэллапа (Gallup 1970). На сегодняшний день установлен факт наличия способности к самоузнаванию у понгид, слонов, дельфинов, касаток и сорок (подробнее см.: Хватов, 2014). Недавно было высказано предположение о наличии данной способности у беспозвоночных - муравьев (Cammaerts, Cammaerts, 2015). Однако са- моузнавание в зеркале является лишь частным аспектом эволюции схемы тела животных. Кроме того, экспериментальную процедуру выявления данной способности трудно применять по отношению к животным, у которых зрительная модальность не является ведущей (например, к большинству млекопитающих; см.: Bekoff, 2001).
В рамках разрабатываемой одним из авторов данной статьи концепции самоотра- жения животных и человека (Хватов, 2014), базирующейся на онтологическом подходе (Барабанщиков, 2002), схема тела рассматривается как один из продуктов процесса самоотражения, а также как филогенетическая основа формирования самосознания. Самоотражение определяется как процесс и как результат отражения субъектом своей внутренней объективной реальности: характеристик своего организма, а также, если речь идет о человеке, своих свойств в качестве участника социальных процессов. Самоотражение рассматривается системно в контексте своей взаимосвязи с отражением внешней среды как два элемента единого психического отражения индивидом акта своего со-бытия с окружающей объективной реальностью.
Авторским коллективом настоящей статьи было дано целостное описание процесса развития самоотражения в ходе филогенеза животных (Хватов, 2010; 2014), а также в рамках филогенеза отдельных крупных таксонов (Хватов и др., 2013b; 2014). Также был проведен ряд экспериментальных исследований особенностей процесса самоотражения и схемы тела у отдельных видов животных (Хватов, 2010; 2011; Хватов, Харитонов, 2012, 2013; Хватов, Харитонов, Соколов, 2014; Хватов и др., 2013a; 2014a; 2014b). В этих исследованиях схема тела животных изучалась в процессе соотнесения ими физических параметров собственного тела с пространственными характеристиками внешних объектов при решении различного рода локомоторных задач, а также при формировании навыков пространственной ориентации.
К настоящему моменту накоплен значительный пласт данных, свидетельствующих о наличии способности формировать навыки пространственной ориентации различной степени сложности у отдельных видов змей, черепах и ящериц, как в естественных, так и в лабораторных условиях (подробнее см.: Burghardt, 1977; Сафаров, 1990). В частности, было установлено, что ящерицы способны запоминать пространственное расположение пищевой приманки, а также укрытий (Brattstrom, 1978; Adler, Phillips, 1985; Day et al., 1999; 2003; Punco, Madragon, 2002; Carazo et al., 2008; Paulissen, 2008; LaDage et al., 2012; Noble et al., 2012). Имеются данные о способности ящериц к переносу приобретенного опыта из одной ситуации в другую при решении дифференцировочной задачи (Leal, Powell, 2012; Wilkinson, Huber, 2012), а также при формировании навыков путем подражания (Noble et al., 2014). Изучены особенности схемы тела королевских змей, а также их способность учитывать увеличение границ собственного тела после кормления (Хватов и др., 2015a; 2015b).
Отдельной вопрос касается того, какая стимуляция имеет ключевое значение для ориентации поведения ящериц. При выработке навыков часто применяют положительное подкрепление (пищу) (Burghardt, 1977), кроме того, существуют данные об ориентации ящериц на визуальные стимулы как в ходе обычного терморегуляционного поведения (Четанов, 2009), так и при выработке пространственных навыков (Wilkinson, Huber, 2012; Clark et al., 2014).
Целью настоящего исследования являлось изучение восприятия границ собственного тела, как одного из ключевых аспектов схемы тела, гигантскими синеязыкими сцинками, а также влияния на данное восприятие объективного увеличения границ тела этих ящериц.
Гипотезы исследования.
1. Сцинки способны научиться учитывать границы собственного тела при взаимодействии с внешними объектами.
2. При изменении границ тела сцинков таким образом, что данные изменения будут препятствовать осуществлению животным ранее выученного поведения, животные модифицируют схему своего тела, что будет выражаться в том, что они сформируют новый навык с учетом объективных изменений границ их тела.
Методика исследования
Животные: 16 сцинков: 8 самцов и 8 самок (половых различий в поведении в рамках эксперимента выявлено не было) гигантских синеязыких сцинков Tiliqua gigas (далее - гигантский сцинк) - взрослые наивные особи, пойманные в дикой природе и наблюдавшиеся в лабораторных условиях около года.
В качестве объектов сцинковые были выбраны именно потому, что для большинства представителей этого семейства (в частности, для гигантских сцинков) характерно роющее поведение, проникновение в различные отверстия в земле, между камнями и корнями деревьев (Mecke et al., 2013). Соответственно, экспериментальная задача являлась экологичной для данного вида.
Животные были разделены на две группы: экспериментальную (8 особей) и контрольную (8 особей).
Оборудование. Экспериментальная установка была организована в прямоугольном вертикальном террариуме (размер террариума: 450x450x620 мм). Внутри террариума располагался проблемный ящик (размер ящика: 150х150х220 мм) (рис. 1). В каждой из боковых стенок ящика располагалось по одному отверстию на высоте 10 мм над уровнем пола.
Диаметр отверстий мог варьироваться тремя способами.
• Малое отверстие (далее - S) диаметром 18 мм, в такое отверстие проходил только кончик морды сцинка;
• Среднее отверстие (далее - M) диаметром 40 мм, в такое отверстие проникала голова сцинка при ее естественных размерах, но не при увеличенных границах;
• Большое отверстие (далее - L) диаметром 70 мм, в такое отверстие проникала голова сцинка и при естественных и при увеличенных границах.
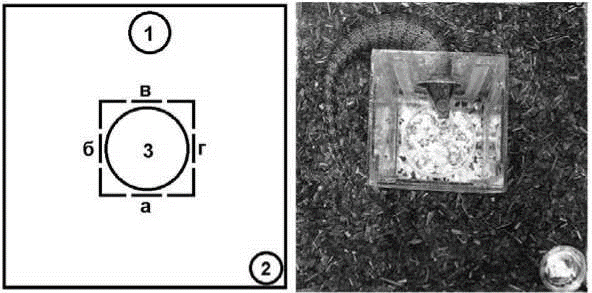
Рис 1. Слева - схема экспериментальной установки (вид сверху): 1 - лампа (над установкой); 2 - поилка; 3 - миска с кормом; а, б, в, г - отверстия в проблемном ящике
Условия в экспериментальном террариуме. В качестве субстрата использовалась смесь: 1/3 - дробленая кора пихты; 1/3 - кокосовое волокно, 1/3 - кокосовая крошка. Субстрат и стенки террариума увлажнялись каждые 2 дня (после проведения экспериментальных проб). Также в террариуме располагалась поилка с водой (см. рис. 1).
Под террариумом располагался термокабель, занимавший половину площади дна террариума. Террариум освещался ультрафиолетовой лампой (UVB200 25ВТ), расположенной вертикально на высоте 50 см (см. рис. 1). Уровень освещения под лампой - 300 люкс. Цикл содержания день/ночь: 12/12 часов. Температура на обогреве: 37о C днем и 24оС ночью. Фоновая температура: 25оС днем и 20оС ночью.
Кормление животного осуществлялось только внутри экспериментальной установки таким образом, что для достижения корма животному необходимо было проникать головой в одно из отверстий. Кормушка (пластиковая подставка диаметром 100 мм на ножках высотой 10 мм) с приманкой помещалась в центр экспериментальной установки в начале каждой пробы.
Процедура эксперимента. Перед началом эксперимента каждый сцинк в течение пяти дней содержался в экспериментальном террариуме для адаптации к экспериментальным условиям. В начале каждой пробы внутрь экспериментальной установки помещалась приманка - корм. В качестве корма использовалась смесь: куриные желудочки, куриная печень, творог, бананы. Время пробы не ограничивалось; проба считалась завершенной, когда сцинк достигал приманки, проникнув в одно из отверстий. Пробы проводились в светлое время суток, между экспериментальными пробами внутри серии выдерживался временной интервал в два дня. По окончании пробы корм изымался из экспериментальной установки. Между пробами отверстия в экспериментальной установке закрывались стеклом, отверстия открывались (сообразно экспериментальному плану) непосредственно перед помещением корма в проблемный ящик. Экспериментальная установка была окружена непрозрачным занавесом для исключения побочного влияния визуальных стимулов на поведение сцинков.
В экспериментальной и контрольной группах проводилось 4 серии по 20 проб каждая (табл. 1).
Таблица 1
Схема организации эксперимента
|
Серии |
Группы |
|
|
Экспериментальная |
Контрольная |
|
|
№ 1 |
Отверстия: № 1 - M; № 2 - S; № 3 - S; № 4 - S. Границы тела - естественные |
|
|
№ 2 |
Отверстия: № 1 - M; № 2 - S; № 3 - L; № 4 - S. Границы тела - измененные |
Отверстия: № 1 - S; № 2 - S; № 3 - M; № 4 - S. Границы тела - естественные |
|
№ 3 |
Отверстия: № 1 - M; № 2 - S; № 3 - M; № 4 - L. Границы тела - измененные |
Отверстия: № 1 - S; № 2 - S; № 3 - S; № 4 - M. Границы тела - естественные |
|
№ 4 |
Отверстия: № 1 - M; № 2 - L; № 3 - M; № 4 - M. Границы тела - измененные |
Отверстия: № 1 - S; № 2 - M; № 3 - S; № 4 - S. Границы тела - естественные |
Серия № 1. Ставилась задача сформировать у сцинков обеих групп навык доставать приманку через отверстие № 1 (другие отверстия имели слишком маленький диаметр). О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества неуспешных попыток достижения приманки через отверстия № 2, 3 и 4.
Серия № 2. Границы тела сцинков экспериментальной выборки увеличивались с помощью крепления на теменную часть головы животного цилиндрического пластикового объекта высотой 12 мм, диаметром 10 мм. Объект крепился непосредственно перед началом каждой пробы и снимался после ее окончания. После увеличения границ тела животные не могли проникать головой в отверстия типа M, т. е. доставать приманку для них было возможно только через отверстия типа L. Ставилась задача: выяснить, смогут ли сцинки при условии увеличения границ их тела до такого размера, что решение прежней задачи ранее выученным способом (получение приманки через отверстие № 1) окажется невозможным, модифицировать собственное поведение для достижения желаемого результата, т. е. найти новый путь через отверстие № 3. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение неуспешных попыток достижения приманки через отверстия № 1, 2 и 4.
В контрольной выборке границы тела сцинков оставались неизменными, однако отверстие № 1, через которое в предыдущей серии у них был сформирован навык достижения приманки, делалось непроницаемым (S), а проницаемым (M) становилось отверстие № 3. Ставилась задача: выявить, смогут ли сцинки при условии неизменности физических параметров тела, но изменении условий внешней среды найти новый путь к приманке. Полученные данные (по количеству проникновений в отверстия и времени решения экспериментальной задачи) сопоставлялись с данными, полученными на испытуемых экспериментальной выборки в этой же серии, чтобы установить, по-разному ли влияет на поведение сцинков ситуация невозможности реализации ранее сформированного навыка, создаваемая изменением параметров тела в экспериментальной группе и изменением параметров внешней среды в контрольной группе.
Серия № 3. Проводилась при условии, что к концу серии № 2 у испытуемых обеих выборок удавалось сформировать новый навык достижения приманки через отверстие № 3.
В экспериментальной группе ставилась задача установить, как быстро и после какого количества ошибок (неуспешных попыток достижения приманки) сцинки смогут сформировать новый навык достижения приманки через отверстие № 4 при условии, что границы тела останутся увеличенными, но размер отверстия № 3 уменьшится до M, т. е. станет непроницаемым для их тела. О формировании навыка должно было свидетельствовать сокращение времени решения задачи, а также уменьшение количества неуспешных попыток достижения приманки через отверстия № 1, 2 и 3.
В контрольной выборке границы тела оставались неизменными, однако отверстие № 3, через которое в предыдущей серии у них был сформирован навык достижения приманки, делалось непроницаемым (S), а проницаемым (M) становилось отверстие № 4. Ставилась задача: выявить, смогут ли сцинки при условии неизменности физических параметров тела, но изменении условий внешней среды, найти новый путь к приманке.
Данные, полученные в экспериментальной и контрольной выборках, сопоставлялись друг с другом.
Серия № 4. Проводилась при условии, что к концу серии № 3 у испытуемых обеих выборок удалось сформировать новый навык достижения приманки через отверстие № 4.
Ставились аналогичные задачи, что и в предыдущей серии для испытуемых обеих выборок, однако теперь приманку можно было достичь через отверстие № 2, а отверстие № 4 становилось непроницаемым.
Данные, полученные в экспериментальной и контрольной выборках, сопоставлялись друг с другом.
Процедура модификации границ тела сцинков. В область лобного щитка на голове сцинков (экспериментальной группы) с помощью цианоакрилата крепился винт из нержавеющей стали шляпкой вниз. В результате данной операции часть винта с резьбой выступала над поверхностью головы на высоту 3 мм. Операция проводилась в соответствии с современными принципами биоэтики, определенными Этическим кодексом Совета международных организаций 1985 г., Европейской конвенцией о защите позвоночных животных, используемых для экспериментов или в иных научных целях 1986 г., и Правилами лабораторной практики Минздрава РФ 2003 г.
К установленному винту в дальнейшем крепился цилиндрический объект высотой 12 мм, диаметром 10 мм (рис. 2). Винт крепился за 5 минут до начала каждой пробы и снимался спустя 5 минут после ее окончания.

Рис 2. Сцинк из экспериментальной группы с закрепленным на голове цилиндрическим объектом
Операция проводилась после линьки животного и за 7 дней до начала экспериментальной серии № 1. Объективным критерием того, что данная операция не оказала влияния на специфику соотнесения животными собственного тела с объектами внешней среды и, соответственно, на их схему тела, должно было являться отсутствие статистических различий в решении задачи между испытуемыми контрольной и экспериментальной выборок по критерию времени и количества проникновений в различные типы отверстий.
Аппаратура. Поведение животных в процессе эксперимента фиксировалось с помощью видеокамеры (Sony HDR-CX405), для фиксации временных интервалов использовался таймер видеозаписи этой камеры. Видеокамера крепилась над экспериментальной установкой - на высоте 750 мм от пола экспериментального террариума.
Математический анализ осуществлялся с помощью программы Statistica 8.
Регистрируемые показатели.
• Время (в секундах) решения экспериментальной задачи в каждой пробе; в обеих группах регистрировалось от момента помещения корма в проблемный ящик до момента достижения сцинком приманки через одно из отверстий.
• Количество попыток проникновения (успешных и неуспешных) в различные типы отверстий отдельно для каждого отверстия в каждой пробе и отдельно для каждого сцинка в каждой пробе во всех сериях, в обеих группах. За одну попытку проникновения считались ситуации, когда сцинк погружал голову в отверстие хотя бы на несколько миллиметров.
Переменные.
Независимые переменные:
• границы тела сцинков: естественные (естественными считались границы тела испытуемых контрольной группы, а также экспериментальной группы в отсутствие крепления цилиндрического объекта), измененные с помощью цилиндрического объекта;
• диаметр отверстий проблемного ящика.
Зависимые переменные:
• время решения экспериментальной задачи в каждой пробе;
• количество успешных/неуспешных попыток проникновения в различные типы отверстий.
Эмпирические критерии подтверждения гипотез исследования.
Гипотеза 1 может считаться подтвержденной в том случае, если за 20 проб серии № 2 испытуемые совершат достоверно меньше неуспешных попыток достижения приманки, нежели за 20 проб серии № 1. Этот факт дал бы основание утверждать, что сцинки не просто научились не проникать в отверстия типа S, располагающиеся в определенных частях экспериментальной установки (в серии № 1), но и в целом научились воспринимать отверстия данного типа как «непроницаемые» даже в случае расположения этих отверстий в других частях экспериментальной установки (в серии № 3). Соответственно, данный факт свидетельствовал бы о том, что сцинки научились учитывать естественные границы собственного тела.
Гипотеза 2 может считаться подтвержденной, во-первых, при условии предварительного подтверждения гипотезы 1, во-вторых, при условии, что во 2-й, 3-й или 4-й сериях будет обнаружено отсутствие достоверных различий по количеству неуспешных попыток достижения приманки, осуществленных испытуемыми контрольной и экспериментальной выборок. Этот факт дал бы основание утверждать, что сцинки экспериментальной выборки научились воспринимать отверстия типа M как «непроницаемые» (аналогично отверстиям типа S). Соответственно, данный факт свидетельствовал бы о том, что сцинки научились учитывать увеличенные границы собственного тела.
Результаты исследования
Серия № 1. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 3). В экспериментальной группе: тест Вилкоксона T=0; Z=2,47; n=8; p<0,05; в контрольной группе: T=0; Z=2,47; n=8; p<0,05. Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 2654 с (SD=787,3), в контрольной группе - 2589 с (SD=520,8). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 89 с (SD=39,2), в контрольной группе - 92 с (SD=25,6). По показателям времени решения экспериментальной задачи (суммарно за все 20 проб) испытуемые экспериментальной и контрольной групп не отличались друг от друга (тест Манна-Уитни U=31,0; n1=8; n2=8; p>0,05).
В экспериментальной и контрольной группах к концу серии сцинки стали чаще совершать попытки проникновения в отверстия № 1 и реже - в отверстия № 2, 3 и 4 (рис. 3). В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 1 составляла 17%, на последних пяти пробах - 100%; х2=116,15; df=3; p<0,01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 1 составляла 19%, на последних пяти пробах - 100%; х2=113,58; df=3; p<0,01. По общим соотношениям количества попыток проникновения в отверстия различных номеров достоверных отличий у испытуемых двух групп не обнаружено (х2=0,37; df=3; p>0,05).
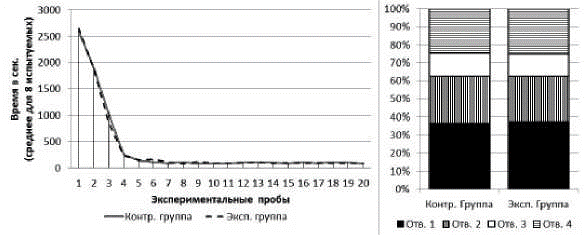
Рис. 3. Слева - кривые научения контрольной и экспериментальной групп в серии № 1.
Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 1
Серия № 2. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 4). В экспериментальной группе: T=0; Z=2,52; n=8; p<0,01; в контрольной группе: T=0; Z=2.52; n=8; p<0.01. Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 715 с (SD=104,8), в контрольной группе - 315 с (SD=33,1). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 97 с (SD=31,4), в контрольной группе - 105 с (SD=33,9). На решение экспериментальной задачи сцинки обеих групп в серии № 2 затратили меньше времени, нежели в серии № 1. В экспериментальной группе: тест Вилкоксона: T=0; Z=2,52; n=8; p<0,05; в контрольной группе: T=0; Z=2,52; n=8; p<0,05. При этом на решение экспериментальной задачи испытуемые экспериментальной группы затратили больше времени (суммарно за все 20 проб), чем испытуемые контрольной группы (U=2; n1=8; n2=8; p<0,01).
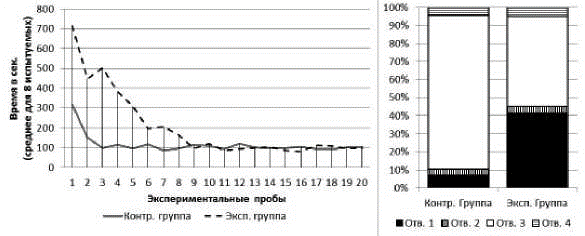
Рис. 4. Слева - кривые научения контрольной и экспериментальной групп в серии № 1. Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 2
В экспериментальной и контрольной группах к концу серии сцинки стали чаще совершать попытки проникновения в отверстия № 3 и реже - в отверстия № 1, 2 и 4. В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 3 составляла 24%, на последних пяти пробах - 100%; х2=78,18; df=3; p<0,01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 3 составляла 59%, на последних пяти пробах - 100%; х2=22.23; df=3; p<0,01 (рис. 4).
У испытуемых обеих выборок доля неуспешных попыток достижения приманки в серии № 2 снизилась в сравнении с серией № 1 (для экспериментальной выборки - х2=10,84; df=1; p<0,01; для контрольной выборки - х2=121,70; df=1; p<0,01). При этом в серии № 2 сцинки экспериментальной группы совершили больше неуспешных попыток, нежели сцинки контрольной группы (х2=62,61; df=1; p<0,01).
Серия № 3. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 5). В экспериментальной группе: T=0; Z=2,52; n=8; p<0,01; в контрольной группе: T=0; Z=2,52; n=8; p<0,01. Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 503 с (SD=65,9), в контрольной группе - 254 с (SD=50,0). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 96 с (SD=34,5), в контрольной группе - 112 с (SD=31,1). На решение экспериментальной задачи сцинки экспериментальной группы в серии № 3 затратили меньше времени, нежели в серии № 2 (тест Вилкоксона: T=1; Z=2,38; n=8; p<0,05). В контрольной группе такого снижения времени не выявлено (тест Вилкоксона: T=18; Z=0; n=8; p>0,05). При этом достоверных различий между испытуемыми разных групп по критерию общего времени решения задачи (суммарно за 20 проб) выявлено не было (U=14; n1=8; n2=8; p>0,05).
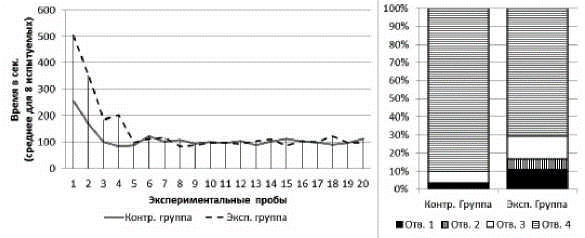
Рис. 5. Слева - кривые научения контрольной и экспериментальной групп в серии № 1. Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 3
В экспериментальной и контрольной группах к концу серии сцинки стали чаще совершать попытки проникновения в отверстия № 4 и реже - в отверстия № 1, 2 и 3. В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 4 составляла 38%, на последних пяти пробах - 100%; х2=55,61; df=3; p<0,01. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 4 составляла 69%, на последних пяти пробах - 100%; х2=17,09; df=3; p<0,01 (рис. 5).
У испытуемых экспериментальной группы доля неуспешных попыток достижения приманки в серии № 3 снизилась в сравнении с серией № 2 (х2=22,31; df=1; p<0,01). У испытуемых контрольной группы такого снижения выявлено не было (х2=1,90; df=1; p>0,05). При этом в серии № 3 сцинки экспериментальной группы совершили больше неуспешных попыток, нежели сцинки контрольной группы (х2=23,26; df=1; p<0,01).
Серия № 4. У испытуемых обеих групп от 1-й к 20-й пробе наблюдалось снижение времени решения экспериментальной задачи (рис. 6). В экспериментальной группе: T=0; Z=2,52; n=8; p<0,01; в контрольной группе: T=0; Z=2,52; n=8; p<0,01. Среднее время решения экспериментальной задачи на 1-й пробе в экспериментальной группе составило 249 с (SD=30.9), в контрольной группе - 266 с (SD=31.4). Среднее время решения экспериментальной задачи на 20-й пробе в экспериментальной группе составило 107 с (SD=31,7), в контрольной группе - 104 с (SD=24,1). Не выявлено различий в количестве времени (суммарно за 20 проб), затраченного испытуемыми обеих выборок, между сериями № 3 и 4 (в экспериментальной группе тест Вилкоксона: T=5; Z=1,82; n=8; p>0,05; в контрольной группе: T=17; Z=0.14; n=8; p>0,05). При этом достоверных различий между испытуемыми разных групп по критерию общего времени решения задачи (суммарно за 20 проб) выявлено не было (U=30,5; n1=8; n2=8; p>0,05).
В экспериментальной и контрольной группах к концу серии сцинки стали чаще совершать попытки проникновения в отверстия № 2 и реже - в отверстия № 1, 3 и 4. В экспериментальной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 2 составляла 73%, на последних пяти пробах - 100%; х2=12,95; df=3; p<0,05. В контрольной группе на первых пяти пробах серии доля попыток проникновения в отверстие № 2 составляла 68%, на последних пяти пробах - 100%; х2=15,94; df=3; p<0,01 (рис. 6).
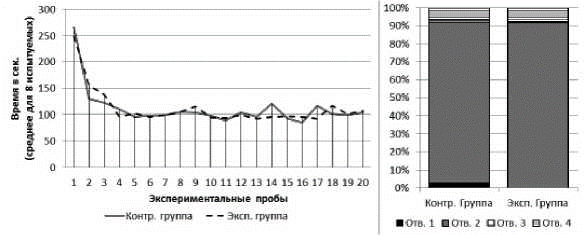
Рис 6. Слева - кривые научения контрольной и экспериментальной групп в серии № 1. Справа - распределения общего количества проникновений в различные отверстия в контрольной и экспериментальной группах в серии № 4
У испытуемых экспериментальной группы доля неуспешных попыток достижения приманки в серии № 3 снизилась в сравнении с серией № 2 (х2=27,35; df=1; p<0,01). У испытуемых контрольной группы такого снижения выявлено не было (х2=0,02; df=1; p>0,05). При этом не было выявлено различий в количестве неуспешных попыток, совершенных испытуемыми экспериментальной и контрольной выборок, в серии № 4 (х2=0,43; df=1; p>0,05).
Обсуждение результатов
Полученные результаты свидетельствуют, что у сцинков обеих групп успешно складывался навык решения экспериментальной задачи. Причем в каждой серии навык формировался заново - как при изменении границ тела, так и при изменении схемы расположения отверстий, о чем свидетельствует факт того, что в начале каждой серии у животных возрастало как время решения экспериментальной задачи, так и количество неуспешных попыток проникновения в непроницаемые отверстия.
Однако мы имеем основание утверждать, что сцинки научились учитывать границы собственного тела при взаимодействии с объектами внешней среды к концу серии № 1. Об этом свидетельствует тот факт, что в серии № 2 сцинки обеих группы совершали меньше неуспешных попыток достижения приманки, нежели в серии № 1. Это означает, что в ходе серии № 1 ящерицы не только выучили схему расположения отверстий в лабиринте, но и то, что отверстие типа S непроницаемо для их головы. В серии № 2, когда схема расположения отверстий изменилась, они перенесли данный опыт в новую ситуацию, в которой ранее проницаемое отверстие стало слишком малым.
У испытуемых контрольной выборки навык сложился (достиг устойчивого состояния) уже к серии № 2, поскольку в последующих сериях не изменялась доля неуспешных попыток достижения приманки, а также общее время решения задачи (суммарно за 20 проб).
У испытуемых же экспериментальной группы увеличение границ тела вызвало, как мы полагаем, большие трудности в дальнейшем решении задачи. Это выражалось в том, что они в сравнении с испытуемыми контрольной группы совершали больше неуспешных попыток достижения приманки и тратили больше времени на решение задачи в сериях № 2 и № 3. Однако эти различия исчезли в серии № 4. Мы объясняем это тем, что к 4-й серии у ящериц экспериментальной выборки завершилась выработка модифицированной схемы тела путем включения в нее инородного цилиндрического объекта. Животные научились учитывать увеличенные границы собственного тела, благодаря чему в последней серии их поведение не отличалось от поведения сцинков контрольной группы.
Таким образом, мы можем заключить, что вторая гипотеза нашего исследования также была подтверждена.
В ранних исследованиях, проводившихся на членистоногих (в частности, тараканах Periplaneta americana (Хватов, 2011)), в ходе которых у животных экспериментальной группы также увеличивали границы тела, а для контрольной группы изменялась схема отверстий в экспериментальной установке, были получены существенно иные результаты. Во-первых, членистоногие, как при изменении границ тела, так и при изменении размера отверстий, демонстрировали одинаковое поведение (по временным параметрам формирования навыка и по соотношению количества попыток проникновения в различные типы отверстий). Во-вторых, членистоногие не переносили ранее приобретенный опыт (как при естественных, так и при измененных границах их тела) из одной серии в другую - в каждой следующей серии навык формировался у них «с нуля»: животные совершали такое же количество неуспешных попыток проникновения в отверстия, слишком маленькие для границ их тела, как и в предыдущей серии.
В аналогичных экспериментах, проводившихся на змеях (Хватов и др., 2015b), было установлено, что эти животные, аналогично членистоногим, не переносят приобретенный опыт о границах своего тела в новую ситуацию, как в случае изменения расположения отверстий в лабиринте, так и в случае увеличения границ их тела.
В исследованиях этого типа, проводившихся на крысах (Хватов и др., 2016), было установлено, что эти млекопитающие способны переносить приобретенный опыт о границах собственного тела в новую ситуация. Кроме того, они, как и сцинки, способны модифицировать схему собственного тела при увеличении границ их тела. Однако, в отличие от сцинков, у которых данный навык модификации схемы тела формируется в течение нескольких экспериментальных серий, у крыс этот навык складывается значительно быстрее - в течение нескольких неуспешных попыток достижения приманки через ранее проницаемое отверстие. Вероятно, у этих двух видов животных модификация схемы тела осуществляется различными механизмами: у сцинков происходит формирование классического навыка, а у крыс происходит своего рода инсайт (Кёлер, 1930).
С позиции разрабатываемой нами концепции самоотражения животных и человека (Хватов, 2010; 2014), вышеизложенные результаты, полученные на представителях различных видов животных, можно проинтерпретировать следующим образом. У сцинков в сравнении с членистоногими обнаруживается большая автономия (и, соответственно, возможность для более гибкой трансформации) схемы собственного тела от схемы внешней среды в рамках целостного психического отражения организмом собственного со-бытия с внешней средой. Это проявляется в том, что животные уже способны модифицировать схему собственного тела путем научения, постепенно обобщая опыт собственного взаимодействия с объектами внешнего мира в различных ситуациях, а также могут переносить опыт о границах собственного тела, приобретенный в одной ситуации, в новую ситуацию. С другой стороны, крысы, а также другие млекопитающие, включая приматов (Herman et al., 2001; Maravita, Iriki, 2004; Johnson-Frey, 2004), способны еще более гибко и быстро приспосабливать схему собственного тела к конкретным условиям внешней среды, а также модифицировать ее при объективном увеличении границ их тела.
Выводы
На основании полученных результатов мы констатируем, что обе гипотезы настоящего исследования были подтверждены. Сцинки способны научаться учитывать границы собственного тела при взаимодействии с объектами окружающей среды. При увеличении естественных границ их тела таким образом, что данные изменения препятствуют осуществлению животным ранее выученного поведения, эти животные модифицируют схему своего тела, что выражается в формировании нового навыка с учетом объективных изменений границ их тела.
[*] Хватов И. А. Кандидат психологических наук, руководитель Центра биопсихологических исследований, Московский институт психоанализа. E-mail: ittkrot1@gmail.com
[†] Соколов А. Ю. Кандидат биологических наук, старший научный сотрудник, Лаборатория-студия «Живая Земля». E-mail: apophis-king@mail.ru
[‡] Харитонов А. Н. Кандидат психологических наук, научный сотрудник, Институт психологии РАН; ведущий научный сотрудник, Центр экспериментальной психологии, Московский государственный психолого-педагогический университет. E-mail: ankhome47@list.ru