Королькова О.А., кандидат психологических наук, старший научный сотрудник, Центр экспериментальной психологии, ФГБОУ ВО МГППУ; преподаватель, кафедра общей психологии, Московский институт психоанализа, Москва, Россия, olga.kurakova@gmail.com
Представления о механизмах восприятия лица человека могут быть существенно дополнены рядом новых данных, полученных с помощью современных методов функционального картирования мозга. В обзоре приводятся результаты исследований, позволяющих уточнить функционирование распределенной мозговой системы, связанной с восприятием лица. Рассматривается проблема того, как соотносятся узнавание человека по лицу и определение его экспрессии. Анализируются механизмы восприятия статических и динамических лиц, а также лица в контексте действия. Подчеркивается необходимость усиления экологической валидности нейрокогнитивных исследований восприятия лица.
Введение
В 1986 г. В. Брюс и Э. Янг предложили функциональную модель восприятия лица человека, включающую ряд обособленных блоков, которые обеспечивают переработку различных типов информации: структурное кодирование изображений лица; узнавание человека по его лицу; распознавание речевой и эмоциональной экспрессии; извлечение из памяти сведений о знакомых людях и др. [Bruce, 1986]. Последовавшие за этим интенсивные нейрокогнитивные исследования позволили соотнести отдельные компоненты данной модели со специфическими паттернами мозговой активации. На основании результатов, полученных к 2000 г., Дж. Хаксби и др. описали распределенную систему мозга, состоящую из базовой части - областей, в которых происходит анализ зрительной информации о лице и его экспрессиях, и расширенной части - областей, связанных с дальнейшей когнитивной переработкой: извлечением семантической информации о знакомых людях, их личностных чертах и отношении к ним; оценкой выраженной на лице эмоции; восприятием направления взора [Haxby, 2000]. Благодаря своему обобщающему характеру, модели Брюс—Янга и Хаксби и др. стали классическими в когнитивной психологии и, с учетом более поздних дополнений, составляют сегодня один из основных подходов к восприятию лица [Haxby, 2011; Ishai, 2008]. Однако со времени публикации этих моделей появился ряд новых данных, которые могут уточнить имеющиеся представления о мозговых механизмах восприятия лица. Предлагаемый обзор включает как краткое описание основных результатов ранних исследований, так и некоторые новые результаты, полученные за последние годы. Акцент сделан на следующих вопросах.
• Как соотносятся узнавание знакомого лица и восприятие его экспрессии?
• Отличается ли восприятие лица в статике и динамике?
• Как воспринимается лицо в контексте действия?
Первая из этих тем поднималась еще Брюс и Янгом 30 лет назад; вторая и третья начали интенсивно исследоваться с помощью нейроимиджинговых методов относительно недавно, и их рассмотрение существенно дополняет классическую модель, опирающуюся прежде всего на данные о восприятии статичных изображений изолированного лица.
Узнавание лица и восприятие экспрессий: независимая переработка или взаимодействие?
Классическая модель предполагает существование двух относительно независимых механизмов: 1) узнавания знакомых лиц, инвариантного относительно изменения выражения лица, направления взора или угла поворота головы; 2) определения экспрессий (эмоциональных и речевых), инвариантного относительно знакомости лица [Bruce, 1986; Haxby, 2000; Hoffman, 2000]. Первый процесс преимущественно связывается с активацией так называемой зоны лиц в нижней затылочной извилине - OFA[I] и зоны лиц в веретеновидной извилине - FFA; второй - с активацией области в задней части верхней височной извилины и борозды - pSTS (рис. 1). Однако более новые данные - как психофизические, так и нейрофизиологические - ставят под сомнение полную независимость этих двух процессов [Calder, 2005]. Рассмотрим некоторые аргументы, которые свидетельствуют как за, так и против гипотезы независимости.
Первый источник данных относительно соотношения двух механизмов составляют случаи нарушений восприятия лица вследствие травм, нейрохирургического вмешательства или врожденных аномалий. Ряд ранних нейропсихологических исследований выявил двойную диссоциацию между узнаванием лица и определением его выражения. В частности, при приобретенной прозопагнозии, развивающейся в результате повреждений в затылочно-височных отделах мозга, нарушается узнавание знакомых лиц, хотя распознавание экспрессии может сохраняться [A case of, 1983; Tranel, 1988]. У других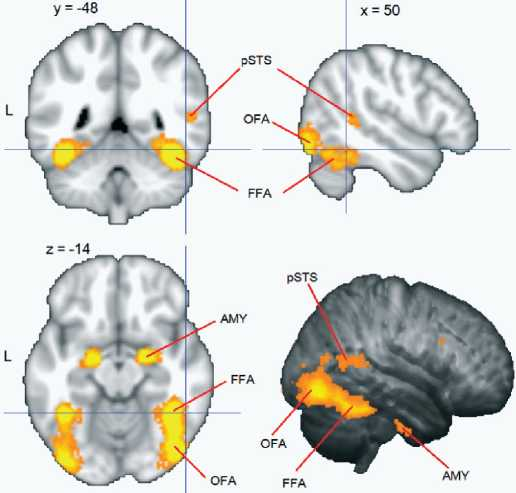
Рис. 1. Области мозга, демонстрирующие избирательную активацию при восприятии изображений лица, представлены на срезах в трех проекциях и в трехмерной реконструкции.
Активация получена по результатам метаанализа 720 исследований, отобранных из базы Neurosynth [Large-scale automated synthesis, 2011] по ключевому слову «face» (http://neurosynth.org/analyses/terms/face/). Отмечены зоны, входящие в ядро распределенной системы: OFA - зона лиц в нижней затылочной извилине; FFA - зона лиц в веретеновидной извилине; pSTS - область в задней части верхней височной извилины и борозды; а также AMY - миндалина, которая входит в расширенную систему и вовлекается в анализ аффективной информации. Координаты срезов соответствуют системе Монреальского неврологического института
пациентов наблюдается противоположная картина: нарушается распознавание экспрессии, но не узнавание человека по его лицу [Face perception after, 1993; Hornak, 1996]. Метаанализ 100 случаев приобретенной прозопагнозии, описанных начиная с 1974 г., показал, что подобные нарушения преимущественно связаны с повреждениями в области OFA и реже - в FFA либо STS [Bouvier, 2005]. Первые исследования прозопагнозии неоднократно подвергались критике с точки зрения используемых методов [Calder, 2005], однако с учетом этой критики за последние годы описано еще несколько случаев, подтверждающих гипотезу независимых механизмов [Are faces special, 2008; Face perception is, 2013; Perceptual and anatomic, 2011]. Они связаны в основном с повреждениями нижней, средней либо передней височной коры или нижней затылочной извилины преимущественно в правом полушарии [Barton, 2008], что также соответствует анатомической модели Хаксби и др., в которой этим зонам отводится ключевая роль в переработке инвариантных характеристик лица. У пациентов с прозопагнозией, имеющих повреждения в OFA, активация в FFA при восприятии лиц была сохранна [Rossion, 2003; The fusiform face, 2006], хотя последняя область при этом не демонстрировала эффекта адаптации — характерного для нормы снижения активации при повторном предъявлении лица одного и того же человека [Abnormal face identity, 2009]. Это свидетельствует о необходимости нормального функционирования области OFA для эффективного распознавания лица. Нарушения восприятия экспрессий при сохранной способности узнавания лиц наблюдаются преимущественно при повреждениях задней части верхней височной борозды (pSTS) и миндалины [Emotional memory and, 2004; Perceptual and anatomic, 2011]. При этом у одного из пациентов с повреждением pSTS были нарушены как распознавание экспрессий, так и узнавание людей с различными экспрессиями, что говорит о возможном взаимодействии двух подсистем [Perceptual and anatomic, 2011]. Однако такой поведенческий эффект может быть связан не только с системой переработки информации о лице, но и с иной системой, обеспечивающей восприятие и переживание эмоций [Calder, 2005], поэтому он не может однозначно трактоваться как доказательство взаимодействия механизмов. В отличие от приобретенных нарушений, врожденная прозопагнозия характеризуется избирательной неспособностью узнавания лиц [Duchaine, 2003; Humphreys, 2007] или экспрессий [Prosopagnosia as an, 2006; Event-related potentials reflect, 2007], возникшей в результате атипичного развития мозга, и обычно не сопровождается другими когнитивными нарушениями [Covert face recognition, 2009; Duchaine, 2007]. Предположительно, такие пациенты могут использовать особые стратегии, позволяющие им в ряде случаев выполнять задания на распознавание и запоминание экспрессий на уровне нормы [Bate, 2015; Humphreys, 2007], поэтому для уточнения особенностей функционирования именно перцептивных механизмов могут потребоваться дальнейшие исследования.
Помимо изучения клинических случаев прозопагнозии, в последнее время широко используется так называемый метод «виртуального пациента», позволяющий с помощью транскраниальной магнитной стимуляции (ТМС) временно инактивировать определенный участок коры мозга у здорового человека и тем самым смоделировать его избирательное повреждение [Pitcher, 2011; TMS Evidence for, 2007]. Методические ограничения ТМС не позволяют воздействовать на зоны, расположенные глубже чем 2-3 см от поверхности коры (в их числе FFA и передняя височная кора), однако направленное воздействие на STS или OFA возможно. Поскольку предполагается, что последняя посылает сигнал в вышележащие области, с помощью данного метода можно исследовать функциональную организацию распределенной системы и эффективность восприятия различных аспектов лица. Показано, что ТМС-инактивация области OFA избирательно нарушает распознавание изображений лиц, но не изображений неживых объектов или тела человека [Triple Dissociation of, 2009], причем это нарушение связано с восприятием изменения отдельных частей лица, но не расстояний между ними [TMS Evidence for, 2007]. По сравнению с контрольным условием, после воздействия на правую OFA активация в FFA (билатерально) на последовательно экспонируемые лица различных натурщиков уменьшалась [Solomon-Harris, 2014], как если бы происходила потеря информации о том, кому принадлежит данное лицо. Однако в отличие от данных, полученных на пациентке с прозопагнозией, которая имела повреждения правой нижней височной извилины и левой веретеновидной извилины [Impaired face discrimination, 2006], активация FFA на разные лица все равно была значимо выше, чем на одинаковые, что предполагает лишь частичное нарушение ее функций вследствие воздействия на OFA. При этом такое воздействие полностью разрушает вклад STS в различение лиц, поскольку активация на разные лица в этой области снижается до уровня активации на повторяющееся лицо. В самой OFA при этом уменьшался эффект адаптации только к экспозиции одинаковых лиц. Что касается pSTS, ее инактивация с помощью ТМС приводила к снижению перцептивного доминирования эмоциональных лиц в условиях бинокулярного соревнования, но не влияла на доминирование нейтральных лиц [Virtual lesion of, 2015], что говорит скорее в пользу различных механизмов их переработки. В других исследованиях, однако, воздействие на STS нарушало восприятие направления взора, но не восприятие экспрессии, на которое влияли только воздействия на область в соматосенсорной коре либо в OFA [Transcranial magnetic stimulation, 2008; Dissociable roles of, 2004]. Последнее может быть связано с селективностью OFA к структурным изменениям частей лица, которые происходят в том числе при изменении экспрессии.
Второй источник данных о независимости либо взаимодействии механизмов переработки двух типов информации - результаты, полученные с помощью метода фМРТ-адаптации. Он основан на том, что по сравнению с экспозицией различающихся стимулов последовательная экспозиция стимулов с одинаковыми характеристиками приводит к снижению активации в зонах, чувствительных к данной характеристике - например, к экспрессии или к личности изображенного человека. При этом одновременные изменения нерелевантной характеристики не меняют активацию данной зоны. Одно из первых исследований продемонстрировало значимую адаптацию к экспрессии в средней части правой STS (mSTS), расположенной кпереди от выделяемой обычно зоны лиц, и на уровне тенденции - адаптацию в правой pSTS; при этом адаптация к личности натурщика наблюдалась в правых pSTS и FFA [fMRI-adaptation reveals dissociable, 2004], тем самым подтверждая гипотезу о разделении функций, хотя по несколько иному анатомическому принципу, чем предполагалось в модели Хаксби и др. Другие исследования также хорошо согласуются с этой моделью, выявляя адаптацию к структурным изменениям лица в области OFA и ряде областей в лобной коре [The correlates of, 2009; Morphing Marilyn into, 2005], а адаптацию к личности - в FFA (билатерально, но преимущественно справа), в нижнелобной коре, в передней части STS (справа), но не в ее задней части [Andrews, 2004; Mazard, 2006; Morphing Marilyn into, 2005]. В области FFA и в миндалине при этом наблюдаются признаки категориального восприятия, поскольку активация при последовательном предъявлении изображений из разных перцептивных категорий выше, чем при демонстрации изображений из одной категории с такими же физическими различиями [The correlates of, 2009; Harris, 2014; Morphing Marilyn into, 2005]. Субъективная степень знакомости лица положительно коррелирует с величиной эффекта фМРТ-адаптации в более передних областях (полюс левой височной доли и правый гиппокамп), которые, вероятно, участвуют в извлечении из памяти семантической информации об известных личностях. Впоследствии адаптация к экспрессиям различных модальностей была продемонстрирована как в STS, так и в FFA [The correlates of, 2009; Top-Down Control of, 2013; Xu, 2010], что говорит о включении обеих областей в анализ изменчивых характеристик лица [Calder, 2005]. Чувствительность различных зон к изменениям экспрессии не обязательно означает, что их функции идентичны. Так, FFA и pSTS могут извлекать из изображений экспрессивных лиц информацию различного рода: о типичности конкретного лица по сравнению с усредненным лицом (FFA) и о его эмоциональной валентности (pSTS) [Said, 2011]. Речь также может идти о взаимной модуляции STS и FFA, при которой их избирательное включение в переработку информации о лице или его экспрессии может дополняться функциональным взаимодействием, обеспечивающим более эффективное распознавание отдельных аспектов лица [The correlates of, 2009]. В обеих областях при этом отмечаются нисходящие влияния задачи или субъективной категоризации. Так, внимание к определенному аспекту лица, которое обеспечивается заданием на распознавание лица или экспрессии, опосредует связь между их обработкой и приводит, в частности, к тому, что активация FFA может быть выше при оценке экспрессий, чем при распознавании лиц [The involvement of, 2005]. В задаче идентификации изображенного человека (одинаковые либо разные натурщики последовательно предъявлялись в каждой пробе) в FFA наблюдается снижение эффекта адаптации не только при изменении личности, но и при изменении экспрессии, выраженной на лице, хотя во втором случае активация повышается не столь существенно [Xu, 2010]. В зонах OFA и pSTS при этом изменений не происходит [Hoffman, 2000]. В другом исследовании в задаче, предполагающей внимание к личности натурщика, адаптация к экспрессии, согласованная с ее перцептивной категоризацией, наблюдалась в FFA и mSTS, а адаптация к личности - в FFA и предклинье. Если же задача предполагала внимание к экспрессии, в обоих случаях (изменений как личности, так и экспрессии) помимо адаптации в этих областях выявлена адаптация в pSTS [The correlates of, 2009]. Адаптация в области OFA, как и в других проведенных исследованиях, не зависела от задачи или субъективного восприятия натурщиков и экспрессий, но была связана только со структурными изменениями лица.
Наконец, третьим источником сведений о соотношении узнавания лиц и экспрессий стали исследования восприятия лица в динамике, которые активно проводятся в последние годы [Dynamic Faces: Insights, 2010]. Вопреки предположениям классической модели, было показано, что динамическая информация может облегчать распознавание как лиц, так и экспрессий [Lander, 2015], в том числе и пациентами с прозопагнозией [Movement cues aid, 2015; Longmore, 2013; Reconstructing dynamic mental, 2015]. Уточненная модель восприятия лица, предложенная Э. О’Тул и коллегами, предполагает, что роль pSTS не сводится к анализу динамической информации об экспрессии, направлении взора и движениях губ при речи, но распространяется также на узнавание знакомых лиц по их характерным движениям [O’Toole, 2002]. FFA, в свою очередь, вовлекается в анализ экспрессий лица и так же, как и pSTS, демонстрирует более высокую активацию на эмоциональные, чем на нейтральные лица [Top-Down Control of, 2013]. Два пути анализа различных характеристик лица - его формы и движения - могут взаимодействовать как при выделении структурной информации, способствующей идентификации человека по его движущемуся лицу, так и при выделении информации о произошедших изменениях в лице по его статическим изображениям [Bernstein, 2015]. Недавнее ТМС-исследование показало, что кратковременное «выключение» OFA снижало ответ как на статические, так и на динамические лица в FFA, тогда как в STS снижался ответ только на статические лица, но не на динамические, что предполагает наличие дополнительных связей с другими областями (в частности, прямые связи STS с прилежащей к ней зоной V5/MT) и еще более раннее разделение путей анализа статической и динамической информации о лице [Pitcher, 2014]. ТМС-инактивация области STS при этом избирательно снижала активацию на динамические, но не на статические лица, подчеркивая ее роль в анализе изменчивых компонентов лица. Эти данные, как и ряд других, предполагают, что два пути переработки информации о лице - вентральный путь анализа структуры лица и латеральный путь анализа его динамики - имеют более высокоуровневые взаимосвязи, чем постулировалось в модели Хаксби и др., либо существует отдельный путь, напрямую соединяющий области в первичной зрительной коре и верхнюю височную извилину [Pitcher, 2014]. Предположительно, движение лица, воспринимаемого в реальной ситуации социального взаимодействия, будет одновременно активировать оба механизма [Lander, 2015].
Рассмотренная совокупность данных о нарушениях восприятия лица и экспрессий свидетельствует скорее в пользу достаточно раннего разделения двух путей переработки информации, оставляя, однако, возможность их последующего взаимодействия. Поскольку повреждения OFA существенно затрудняют распознавание лиц, но значительно меньше влияют на распознавание экспрессий, можно говорить о частичной диссоциации двух механизмов. Вместе с тем, и FFA, и STS, предположительно получающие входной сигнал не только от ипсилатеральной OFA, но и от других зон распределенной системы, могут участвовать в анализе как экспрессии, так и личности изображенного человека, обеспечивая обработку статических и динамических признаков его лица соответственно. Нисходящие влияния внимания и стоящей перед наблюдателем задачи в свою очередь способны модулировать активацию и интенсивность взаимодействия между областями. В отличие от классической модели, результаты новых исследований предполагают, что оба типа данных - и статические, и динамические - используются для более эффективного решения перцептивных задач относительно узнавания натурщика по его лицу и определения его эмоционального состояния.
Восприятие лица в статике и динамике: один механизм или разные?
В предыдущем разделе был приведен ряд данных в пользу взаимодействия механизмов узнавания лица и идентификации его экспрессии, полученных с использованием динамических стимулов. В связи с тем, что лицо в своем естественном движении является более экологически валидным объектом восприятия, чем статичная фотография [Enhanced neural activity, 2004], изучение лежащих в основе этого процесса мозговых механизмов представляется актуальной задачей, решению которой посвящено возрастающее число работ. В ряде исследований были сделаны попытки ответить на вопрос, обеспечивается ли восприятие экологически валидных динамических экспрессий особым механизмом, не сводимым к механизмам восприятия статичных лиц. Сопоставление мозговой активации в ответ на экспозицию статических и динамических лиц показывает, что последние действительно вызывают более стабильный и интенсивный ответ во всех ключевых зонах системы восприятия лица [Fox, 2009; Differential selectivity for, 2011]. При этом наибольшее увеличение активации наблюдалось в области pSTS, которая играет ключевую роль в восприятии биологического движения, но которую зачастую оказывается невозможным выявить у отдельного испытуемого при использовании в качестве стимульного материала статических изображений [Andrews, 2004]. Вместе с тем, роль движения в зрительном анализе лица не ограничивается интеграцией зон, входящих в ядро распределенной мозговой системы. При восприятии динамических лиц усиливается активация в парагиппокампальной извилине и миндалине, в затылочной и орбитофронтальной коре [Differential selectivity for, 2011; Fox, 2009; Pitcher, 2014; What the Human, 2013; Schultz, 2009]. В анализ динамической информации вовлекаются зоны, которые практически не демонстрировали активации при восприятии статических изображений: область в верхней височной извилине более кпереди от pSTS, а также в нижней лобной извилине [Fox, 2009; Differential selectivity for, 2011]. Последнюю связывают не только с анализом лиц, но и с восприятием и имитацией действий других людей. Анализ функциональных связей показал, что восприятие реалистичных динамических экспрессий лица усиливает взаимосвязи между областями ранней зрительной обработки в затылочной коре и STS, а также между STS и миндалиной и нижней лобной извилиной [Dynamic Faces: Insights, 2010], в полном соответствии с предположением о наличии обратных связей между этими областями [Ishai, 2008]. Между pSTS и более передней частью STS, а также областями в лобной коре имеются и прочные анатомические связи [Explicating the Face, 2013; White-Matter Connectivity between, 2012], что также подтверждает предположение о наличии особого пути переработки динамической информации о лице, ключевой областью которого является STS.
При восприятии эмоциональных экспрессий лица, которые по самой своей природе являются динамическими, движение также приводит к более широкой активации зон мозга. По сравнению со статическими лицами, динамическая экспрессия радости вызывает большую активацию в клиновидной и лингвальной извилинах, зоне V5, связанной с восприятием движения, и STS, в нижней височной (включая FFA) и медиальной фронтальной коре [Dissociable Neural Pathways, 2003; Trautmann, 2009]; экспрессия отвращения - в нижней лобной извилине, миндалине, FFA и STS, [Trautmann, 2009]; экспрессия гнева - в STS, V5 и в области, прилежащей к миндалине [Dissociable Neural Pathways, 2003]. Различные особенности динамики экспрессий также можно дифференцировать по активации мозга: в случае усиления выражаемой эмоции страха по сравнению с ее ослаблением наблюдается изменение активации в левой FFA [Reinl, 2014] и в миндалине [Enhanced neural activity, 2004], а при инверсии видеозаписей экспрессии во времени - в правой pSTS [Reinl, 2014]. Активация в STS выше при экспозиции реалистичных видеозаписей лица натурщика по сравнению с динамическими стимулами, инвертированными в пространстве (поворот на 180°) [Girges, 2016] или со стимулами с перемешанным порядком кадров, а частота кадров, с которой демонстрируется видеозапись, коррелирует с активацией областей, связанных с восприятием как лица, так и движения [What the Human, 2013]. Недавнее МЭГ-исследование восприятия динамических экспрессий радости и гнева, по сравнению с динамическими рандомизированными паттернами, позволило выявить не только структурные, но и временные характеристики процесса восприятия: в интервале 150-200 мс, содержащем специфический для лица компонент M170, возрастает активация в FFA и STS, тогда как на более поздних этапах переработки информации (250-400 мс от начала экспозиции лица) наблюдается как активация в FFA и STS, которая может быть связана с более поздними стадиями когнитивной переработки, так и специфическая активация в нижней лобной коре [Sato, 2015]. Анализ динамики связей между данными областями показал, что наиболее точно наблюдаемую активацию описывает модель, включающая прямые и обратные связи между зонами, организованные в двух направлениях: от зрительной коры через FFA и STS к нижней лобной коре и от зрительной коры через зону V5 и STS. Согласно выдвинутому авторами данного исследования предположению, после начальных этапов зрительной и когнитивной обработки информация об экспрессии лица может использоваться в виде моторной репрезентации для подготовки к дальнейшему действию.
В целом, исследования динамических экспрессий говорят в пользу того, что мозговые механизмы, обеспечивающие восприятие движения лица (прежде всего STS), не сводятся к механизмам восприятия статических лиц, задействуют большее число областей мозга и участвуют в узнавании как лиц, так и экспрессий [Bate, 2015]. Представляется, что STS является ключевой зоной латерального пути анализа лицевого движения и участвует в интеграции информации о форме и движении динамических лицевых стимулов на достаточно ранней стадии переработки.
Восприятие лица в контексте: какие механизмы обеспечивают коммуникацию «лицом к лицу»?
Как уже отмечалось ранее, классические модели переработки информации о лице основываются преимущественно на исследованиях восприятия изображений статичного изолированного лица. Однако в реальной жизни за пределами лаборатории мы редко сталкиваемся с такими ситуациями. Напротив, мы наблюдаем и оцениваем лица, включенные в самый разнообразный контекст, который может оказывать существенное влияние на их восприятие [Wieser, 2012]. Контекст может содержать как дополнительную информацию о действиях, жестах, намерениях и речи изображенного человека, так и сведения о ситуации, в которую он включен. Наконец, в качестве еще одного источника контекста, связанного с самим наблюдателем, можно рассматривать его установки, прошлый опыт и индивидуальные особенности. Полученные за последнее время данные о том, какие структуры мозга и их функциональные взаимодействия обеспечивают анализ лица, включенного в контекст, позволяют приблизиться к пониманию механизмов процесса взаимодействия «лицом к лицу» в повседневной жизни.
При восприятии эмоциональных экспрессий лица одним из контекстных признаков, увеличивающих точность оценки выраженных эмоций, служит направление взора натурщика. В частности, когда взгляд натурщика направлен вперед, динамическая экспрессия гнева средней интенсивности воспринимается как сильнее выраженная и более релевантная для наблюдателя, а активация в миндалине, FFA и медиальной префронтальной коре увеличивается, по сравнению с восприятием лица, взгляд которого направлен в сторону [N’Diaye, 2009; Effects of gaze, 2014]. В случае экспрессии страха, демонстрируемой на короткое время или обладающей низкой интенсивностью, напротив, лицо с отведенным в сторону взором сильнее активирует данные области мозга, а эмоция воспринимается как более интенсивная и релевантная [Amygdala responses to, 2012; N’Diaye, 2009]. Вместе с тем, имеются данные о том, что в анализе направления взора и валентности экспрессии лица участвуют различные участки миндалины, в связи с чем вопрос о взаимодействии механизмов остается открытым [Effects of gaze, 2014].
Еще один важный признак, по которому мы судим о намерениях собеседника и который используем для предвосхищения его дальнейших действий по отношению к нам, - его жесты, поза и экспрессия тела. В отличие от исследований лица, изучение экспрессий тела является относительно новым предметом для социальной и аффективной нейронауки [Standing up for, 2010]. Если два источника данных об экспрессии конгруэнтны, эффективность и скорость анализа эмоционального состояния коммуниканта существенно повышается, а ранняя интеграция информации отражается в электрической активности мозга, тогда как при рассогласованиях экспрессии лица и тела наблюдатель совершает больше ошибок в распознавании выражений лица [Meeren, 2005]. Распределенные системы, связанные с анализом лица и тела, частично пересекаются (в частности, обе включают миндалину, верхнюю височную борозду и веретеновидную извилину [Similarities and differences, 2011; Peelen, 2005]), однако имеется и ряд специфических для восприятия тела областей. В частности, по сравнению с динамическими лицами, динамические изображения тела, выражающего состояния страха либо гнева, сильнее активируют экстастриарную область, связанную с восприятием тела человека (EBA), в затылочной коре, STS и область в височно-теменной связке (которая, согласно ряду исследований, включена в анализ социальных действий); миндалина сильнее активируется на изображения экспрессий лица, чем тела; веретеновидная извилина в целом демонстрирует одинаковую активацию как на лица, так и на тела [Similarities and differences, 2011], однако за их кодирование отвечают различные нейронные популяции [Schwarzlose, 2005]. Специализация областей в затылочной коре, включенных в перцептивный анализ лица (OFA) и тела (EBA), происходит на достаточно ранних стадиях распознавания изображений. С помощью ТМС-инактивации OFA либо EBA удалось избирательно разрушить восприятие лиц либо тел соответственно при воздействии через 100/110 мс после появления стимульного изображения, однако более раннее воздействие (через 40/50 мс) не приводило к такой избирательности [Two critical and, 2012]. С другой стороны, фМРТ-адаптация к изображениям всего тела человека выявила снижение активации в участках веретеновидной извилины и экстрастриарной коры, которое не сводится к сумме эффектов, полученных при адаптации к изображению только лица или только тела, что говорит в пользу не только дифференциации, но и интеграции зрительной информации о теле и лице [Schmalzl, 2012].
Жесты и действия также могут передавать информацию об эмоциональном состоянии человека, которая анализируется как дополнительный контекстный признак, согласованный либо не согласованный с выражением его лица [Yang, 2015]. Восприятие жестов активирует перцептивную и моторную системы, необходимые для распознавания действия, а также эмоциональную систему для анализа намерений и состояния собеседника. При восприятии эмоционального лица в контексте действия активируются области в STS и нижней лобной коре, связанные с анализом движения, подготовкой моторного ответа и предвосхищением действий натурщика [Binding action and, 2013].
Обобщая исследования контекстных эффектов, М. Визер и Т. Брош предложили ряд дополнений в классическую модель Хаксби, согласно которым различные виды контекста могут влиять на разные стадии анализа лицевой информации [Wieser, 2012]. В областях, составляющих ядро распределенной системы, интегрируется информация о дополнительных признаках, извлекаемых из самого лица - направлении взора, динамики экспрессии, а также расовых признаках коммуниканта. Области, входящие в расширенную систему, а также внешние по отношению к лицевой системе зоны, с помощью обратной связи модулируют процессы в ядре системы. Так, внешний контекст предположительно влияет на восприятие лица посредством активации в эмоциональной системе мозга (в частности, в миндалине, островковой коре и стриатуме), которая благодаря системе прямых и обратных связей между миндалиной и зонами ядра лицевой системы усиливает взаимодействие восприятия экспрессий и аффективного контекста, тем самым обеспечивая формирование целостного перцептивного образа. Аналогичные влияния рассмотрены и для других типов контекста. Вероятно, они наблюдаются на всех стадиях процесса анализа лицевой информации, начиная с достаточно ранних, а интеграция сведений, получаемых в том числе и от разных модальностей, является автоматизированной [Beyond the face, 2006; Two critical and, 2012]. Данная распределенная система предположительно обеспечивает сложный процесс восприятия лиц в контексте ситуаций, встречающихся нам в повседневной жизни.
Заключение
В настоящем обзоре был рассмотрен ряд нейрои- миджинговых данных, полученных за последнее время и позволяющих уточнить представления о мозговых механизмах восприятия лица человека. Применение новых методов исследования и анализа данных сделало возможным более детальное изучение строения и функционирования распределенной системы восприятия лица, которая была первоначально описана Брюс и Янгом [Bruce, 1986], Хаксби и др. [Haxby, 2000]. В частности, по результатам последних исследований, в рассматриваемой системе необходимо выделять особый путь анализа динамической информации о лице, который начинается в первичной зрительной коре и в значительной степени отделен от вентрального пути, проходящего через области, связанные с обработкой инвариантных характеристик лица. Динамическая информация может использоваться не только для распознавания изменчивых характеристик лица, как предполагалось ранее, но и для узнавания человека по его характерной мимике, что обеспечивается функциональным взаимодействием различных механизмов. Вероятно, можно говорить о том, что наш мозг «настроен» на анализ движущихся, меняющихся лиц, включенных в контекст и сопровождаемых действиями, взглядом и жестами. Дальнейшие исследования, таким образом, должны быть направлены на более детальное изучение механизмов восприятия экологически валидного, «живого» лица в естественной ситуации коммуникации.
За пределами настоящего обзора остались многочисленные проводимые с помощью ЭЭГ и МЭГ исследования динамики функционирования системы лица, которые также вносят дополнительные уточнения в рабочую модель данного процесса и несомненно заслуживают отдельного рассмотрения. Мы также преимущественно опирались на работы, проведенные на здоровых взрослых респондентах (за исключением анализа случаев прозопагнозии). Однако особенности восприятия лица и его экспрессий также активно изучаются на выборках с различными нарушениями, такими как расстройства аутистического спектра, депрессивные расстройства, шизофрения и др. Мы не анализировали данные, полученные на приматах и представителях других биологических видов, в том числе путем прямой регистрации активности отдельных нейронов, а также на детских и подростковых выборках. Вместе с тем, рассмотренные в данном обзоре работы позволяют составить представление о механизмах восприятия лица и выявить основные тенденции дальнейших исследований.
Благодарности
Работа выполнена при поддержке Российского гуманитарного научного фонда (проект № 15-36-01281 «Структура восприятия динамических экспрессий лица»)
[I] Для краткости используются устоявшиеся англоязычные аббревиатуры: OFA - occipital face area (зона лиц в нижней затылочной извилине); FFA - fusiform face area (зона лиц в веретеновидной извилине); pSTS - posterior superior temporal sulcus (задняя часть верхней височной борозды).