Ахроматическое зрение, по определению, не чувствительно к цветовой составляющей сигнала, а выделяет только его яркость, физическим аналогом которой является интенсивность светового излучения. Вопрос о нейронных механизмах восприятия интенсивности света остается дискуссионным.
Традиционно нервный код интенсивности света рассматривается как соответствие между величиной яркости стимула и такими характеристиками работы нейронов, как частота импульсной активности, величина межимпульсных интервалов, распределение потенциалов действия во времени и т. п. (см., например: Батуев, Куликов, 1983). Существенным в данном случае является не тип реакции, а то, что при любой ее разновидности для восприятия интенсивности света достаточно участия одной, не разлагаемой на самостоятельные компоненты нейрональной системы. Принципиально другой подход связывает восприятие интенсивности света с активностью не одной, а двух независимых нейрональных систем – «яркостной» и «темновой», отвечающих на изменение освещенности одновременно, но противоположным образом (Heggelund, 1974; Фомин, Соколов, Вайткявичус, 1979; Izmailov, Sokolov, 1991). Так, осветление стимула приводит к возбуждению яркостного и торможению темнового каналов, тогда как его потемнение вызывает обратный эффект. При подобном способе реагирования параметры ответов каждой отдельной системы утрачивают свое значение и интенсивность света однозначно связывается только с cоотношением одновременной активности яркостных и темновых базисных элементов. Если отразить характер работы этих элементов геометрически, то получится следующее. Cенсорный образ интенсивности света Si будет представлен вектором X1iX2i в двумерной ортогональной системе координат (X1 – числовая ось активности яркостной системы, а X2 – числовая ось активности темновой системы). Воспринимаемое изменение интенсивности излучения с Si на Sj ведет к изменению координат и формированию нового вектора X1jX2j. В соответствии с этим математическим представлением нервное преобразование, при котором определенному значению интенсивности излучения соответствует определенное соотношение активности яркостной и темновой нейронных систем, получило название векторного кодирования интенсивности света (Соколов, 2003).
Принцип векторного кодирования интенсивности света подтвержден экспериментально при изучении ахроматического зрения человека, рыбы, лягушки и виноградной улитки (Измайлов, Соколов, 1984; Зимачев и др., 1986; Черноризов 1999; Измайлов, Исайчев, Шехтер, 1998). Как психофизические, так и нейрофизиологические результаты этих исследований, обработанные едиными для всех использованных объектов методами, отражены в наборе апостериорных математических моделей. Эти модели, демонстрирующие в геометрической форме стержневые механизмы восприятия интенсивности света различными животными и человеком, во всех случаях представляют собой кривую, которая с высокой степенью точности описывается уравнением окружности, т. е. двухмерной сферы. Таким образом, и у виноградной улитки, и у лягушки, и у рыбы, и у человека модели, отражающие нейрофизиологический базис восприятия интенсивности света, неизменно двухмерны.
Принципиальная идентичность опорных механизмов восприятия интенсивности света у человека и животных во многом определяет значимость исследований клеточных механизмов ахроматического зрения на упрощенных объектах. Особенно привлекательна в этом отношении виноградная улитка: зрительная система данного моллюска естественным образом упрощает световой сигнал, поскольку, в силу неспособности к цветоразличению, воспринимает только яркостную составляющую излучения. Причиной цветовой слепоты виноградной улитки является то, что все ее фоторецепторы содержат одинаковый фотопигмент. Об этом свидетельствует кривая спектральной чувствительности глаза виноградной улитки, имеющая единственный пик в области 475 нм (Von Berg, Shneider, 1972; Черноризов и др., 1992) и близкая к кривой родопсина на номограмме Дартналла (Dartnall, 1953). Таким образом, являясь aхроматом по своей природе, виноградная улитка представляет собой идеальную биологическую модель для изучения механизмов кодирования интенсивности света.
Редукции зрительной функции соответствует упрощенная морфология сетчатки виноградной улитки. Тщательное исследование ультраструктуры глаза Helix pomatia (виноградной улитки) (Зайцева, 1992) и Helix asperca (ближайшего родственника Helix pomatia) (Eakin, Brandenburger, 1967) обнаружило три типа элементов: фоторецепторы, опорные и нервные клетки. Поскольку опорные клетки являются вспомогательными, а нейроны немногочисленны (по данным Brandenburger – 12 штук в основании ретины), основную информационную функцию выполняют фоторецепторы. Эти фоторецепторы типично рабдомерные и несут микровиллы, в которых содержатся светочувствительные гранулы. Помимо клеточного тела, фоторецепторы имеют длинные аксоны, обладающие спайк-генерирующим механизмом. Эти аксоны образуют оптический нерв и в его составе уходят в ЦНС. Таким образом, фоторецепторы виноградной улитки являются элементами, совмещающими собственно рецепторную функцию и функцию первичного сенсорного нейрона.
Электрофизиологическое исследование суммарной активности сетчатки виноградной улитки показало, что ее электроретинограмма состоит из начального быстрого и последующего медленного деполяризационных компонентов. Медленная составляющая ЭРГ пассивно угасает с окончанием действия стимула без заметного «off» эффекта (Gillary, 1970; Шехтер, Зимачев, Аракелов, 1992). Однако несмотря на то, что темновая off-система в суммарной активности сетчатки виноградной улитки в явном виде не проявляется, двухмерность геометрической модели восприятия интенсивности света у этого моллюска показана при использовании в качестве меры яркостного различия именно амплитуды ЭРГ (Измайлов, Исайчев, Шехтер, 1998). На этом основании можно предположить, что, несмотря на видимое отсутствие «off»-компонента в ЭРГ, у виноградной улитки «темновой» и «световой» каналы восприятия интенсивности света начинают оформляться уже на уровне однослойной фоторецепторной сетчатки. Необходимая для этого «специализация» фоторецепторов по знаку реагирования на свет может быть обусловлена как их эндогенными свойствами, так и сетевыми взаимодействиями. О возможности таких взаимодействий говорит наличие аксональных терминалей фоторецепторов, которые подходят к мелким дендроподобным отросткам нейронов, ветвящимся по периферии сетчатки среди аксонов рецепторных клеток (Зайцева, 1992). «Эндогенная» и «сетевая» гипотезы не противоречивы и могут дополнять друг друга.
Допущение о существовании в однослойной сетчатке виноградной улитки фоторецепторов, характеризующихся эндогенными противоположно направленными ответами на освещение, требует электрофизиологического обследования этих фоторецепторов в условиях полной их изоляции от синаптических влияний. Результаты такой проверки представлены в данной работе.
Методика
Глаза виноградной улитки расположены на вершинах задних головных щупалец. При типичном камерном строении они имеют роговицу, хрусталик, стекловидное тело и одно-слойную сетчатку. При подготовке препарата сначала выделялся глазной бокал, затем через круговой надрез по его краям удалялся хрусталик и, наконец, ретина полностью освобождалась, отсекаясь от основания оптического нерва. В таком виде она инкубировалась в трипсине при температуре 37,5° в течение 17–20 мин. Процедура энзиматической обработки сетчатки виноградной улитки позволяла получить большое количество полностью изолированных фоторецепторов, часть которых переносилась в экспериментальную камеру, содержащуюся при комнатной температуре и наполненную проточным физиологическим раствором для беспозвоночных. Пассивное пребывание фоторецепторов в протоке в течение 15 мин обеспечивало их промывку. После этого на фоне темновой адаптации внутриклеточно регистрировались электрические реакции отдельных фоторецепторов, возникающие на освещение. Для стимуляции использовались вспышки белого цвета стандартной яркости – 25 кд/м2 . Время действия светового сигнала варьировало от 10 мс до 10 с в разных экспериментах.
Ионная избирательность светозависимых каналов изолированных фоторецепторов виноградной улитки контролировалась с помощью поочередной замены через проток физиологических растворов, омывающих препарат. В этой процедуре наряду с нормальным физиологическим раствором использовались растворы, лишенные либо ионов натрия, либо кальция, либо обоих этих ионов и сбалансированные трис-ионами.
Результаты
 При обследовании полностью изолированных фоторецепторов виноградной улитки было обнаружено, что по знаку электрофизиологического ответа на освещение они могут быть разделены на два типа: реакция одних состояла в деполяризационном, а других – в гиперполяризационном сдвиге мембранного потенциала. Пример противоположного знака реагирования на свет фоторецепторов разных типов представлен на рис. 1. Фоторецепторы, деполяризующиеся при освещении, характеризуются следующим: на фоне темновой адаптации их потенциал покоя колеблется в пределах от –27 мВ до –40 мВ в разных фоторецепторах. Световая вспышка вызывает рецепторный ответ в виде деполяризационного смещения МП с начальным пиком и последующим замедленным спадом. Фоторецепторы, отнесенные ко второму типу, в отличие от клеток, деполяризующихся при освещении, отвечают на световой стимул продолжительной гиперполяризационной волной, которой предшествует кратковременное и низкоамплитудное отклонение МП в сторону деполяризации. Потенциал покоя фоторецепторов, гиперполяризующихся при освещении, на фоне темновой адаптации составляет от –45 мВ до –56 мВ в разных фоторецепторах. Подобную реакцию продемонстрировали только 28 % клеток от общего числа обследованных элементов.
При обследовании полностью изолированных фоторецепторов виноградной улитки было обнаружено, что по знаку электрофизиологического ответа на освещение они могут быть разделены на два типа: реакция одних состояла в деполяризационном, а других – в гиперполяризационном сдвиге мембранного потенциала. Пример противоположного знака реагирования на свет фоторецепторов разных типов представлен на рис. 1. Фоторецепторы, деполяризующиеся при освещении, характеризуются следующим: на фоне темновой адаптации их потенциал покоя колеблется в пределах от –27 мВ до –40 мВ в разных фоторецепторах. Световая вспышка вызывает рецепторный ответ в виде деполяризационного смещения МП с начальным пиком и последующим замедленным спадом. Фоторецепторы, отнесенные ко второму типу, в отличие от клеток, деполяризующихся при освещении, отвечают на световой стимул продолжительной гиперполяризационной волной, которой предшествует кратковременное и низкоамплитудное отклонение МП в сторону деполяризации. Потенциал покоя фоторецепторов, гиперполяризующихся при освещении, на фоне темновой адаптации составляет от –45 мВ до –56 мВ в разных фоторецепторах. Подобную реакцию продемонстрировали только 28 % клеток от общего числа обследованных элементов.
Световой фон или предшествующее действию тестового стимула освещение уменьшают, вплоть до полного подавления, ответы фоторецепторов обоих типов. Так, предварительное 10-минутное непрерывное освещение приводит к полному исчезновению фоторецепторного ответа как деполяризационного, так и гиперполяризационного типа. Восстановление световой чувствительности фоторецепторов обоих типов достигается в ходе темновой адаптации. Динамика этого процесса контролировалась при использовании тестовой вспышки, подаваемой в ходе темновой адаптации с 5-минутным интервалом. Интенсивность ответной реакции фоторецептора определялась по величине амплитуды начального пика деполяризации (для деполяризующихся рецепторов) или амплитуды волны гиперполяризации (для гиперполяризующихся рецепторов). Во время темновой адаптации чувствительность как деполяризующихся, так и гиперполяризующихся фоторецепторов постепенно возрастала, о чем свидетельствует увеличение амплитуды их реакции на стационарную вспышку. Полное восстановление активности фоторецепторов достигалось после их 18–25-минутного пребывания в темноте.
Следующая серия экспериментов была посвящена определению ионной избирательности светозависимых каналов изолированных фоторецепторов виноградной улитки. Известно, что основными участниками светоуправляемой проводимости фоторецепторов как позвоночных, так и безпозвоночных животных являются, как правило, ионы Na+ и Ca2+ (Бызов, 1971). Поэтому в механизмах генерации как деполяризационного, так и гиперполяризационного фоторецепторных потенциалов виноградной улитки выявлялась роль именно этих ионов. Для этого производилась регистрация электрической активности фото-рецепторов обоих типов при поочередном удалении из инкубационной среды Na+ или Ca2+ (с заменой на трис-ион), а также контролировалось последующее восстановление ответов в нормальном физиологическом растворе после каждой процедуры. Концентрация Na+ и Ca2+ быстро менялась путем замены физиологических растворов через проток.
На рис. 2 демонстрируется влияние ионов Na+ и Ca2+ на ответы фоторецепторов, де-поляризующихся при освещении. В данных клетках отсутствие Na+ во внеклеточной среде приводит к значительной редукции ответа, не вызывая, однако, его полного исчезновения. Пример этого эффекта представлен на рис. 2,А, где видно, что амплитуда рецепторного потенциала при исходном значении 14 мВ сократилась до 3,5 мВ, когда препарат содержался в безнатриевой среде. Дальнейшего падения ответа не зафиксировано, поскольку в таком усеченном виде реакция оставалась и через 12 мин после выведения Na+ из экзогенного раствора. Восстановление полноценного рецепторного потенциала клетки, деполяризующейся при освещении, достигалось «отмыванием» препарата при замене безнатриевого раствора на нормальный. Поскольку отсутствие натрия в инкубационной среде подавило рецепторный ответ не полностью, проверялось действие еще одного, наиболее вероятного участника генерации рецепторного потенциала – ионов Ca2+. Эффект при удалении внеклеточного Ca2+, демонстрируемый на рис. 2,Б, также состоял в том, что амплитуда деполяризационного ответа уменьшилась, но полностью не исчезла. Изолированный фоторецептор, деполяризующийся при освещении, полностью терял эту способность только тогда, когда раствор вокруг него заменялся на среду, не содержащую одновременно ни ионов Na+, ни ионов Ca2+ (рис. 2,В).
Рис. 2. Влияние удаления внеклеточного натрия и кальция на ответ фоторецептора виноградной улитки, деполяризующегося при освещении:
А – Влияние концентрации внеклеточного Na+: а – фоновый ответ фоторецептора на освещение в нормальном растворе; б – ответ на тот же стимул после 12 минут содержания фоторецептора в безнатриевой среде; в – восстановление световой чувствительности фоторецептора после замены безнатриевого раствора на нормальный;
Б – Влияние концентрации внеклеточного Ca2+: г – фоновый ответ на освещение того же фото-рецептора в нормальном физиологическом растворе; д – ответ на освещение после 6-минутной инкубации клетки в бескальциевом растворе; е – восстановление исходного ответа после замены бескальциевого раствора на нормальный;
В – Влияние одновременного удаления внеклеточного Na+ и Ca2+: ж – фоновый ответ на освещение; з – ответ на освещение в среде, не содержащей ионов натрия и альция через 5 минут после замены раствора; и – восстановление утраченного ответа в нормальном физиологическом растворе. Калибровка 10 мВ, 10 с.
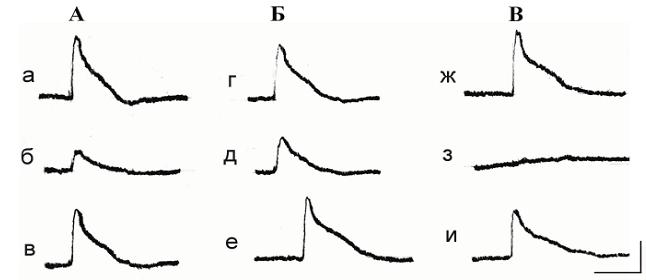
Рис. 3. Влияние удаления внеклеточного натрия и кальция на ответ фоторецептора виноградной улитки, гиперполяризующегося при освещении.
А – Влияние концентрации внеклеточного Na+: а – фоновый ответ фоторецептора на освещение в нормальном растворе; б – ответ на тот же стимул после 3 минут содержания фоторецептора в безнатриевой среде; в – восстановление световой чувствительности фоторецептора после замены безнатриевого раствора на нормальный;
Б – Влияние концентрации внеклеточного Ca2+ : г – ответ на освещение того же фоторецептора в нормальном физиологическом растворе; д – ответ на освещение после 4-минутной инкубации клетки в бескальциевом растворе; е –восстановление исходного ответа после замены бескальциевого раствора на нормальный. Калибровка 10 мВ, 10 с.
 По аналогичной схеме проводились эксперименты, касающиеся ионной избирательности светозависимых каналов фоторецепторов другого типа, в отличие от первых, не де-поляризующихся, а гиперполяризующихся при освещении. В этих опытах, как и в первом случае, контролировалось влияние ионов Na+ и Ca2+ на состояние фоторецепторного ответа, включающего слабую кратковременную начальную деполяризацию и следующую затем более продолжительную и глубокую гиперполяризацию. Влияние на исследуемый ответ содержания экзогенного Na+ проиллюстрировано на рис. 3,А. Из приведенных записей видно, что выведение Na+ из омывающего клетку раствора, не влияя на начальную деполяризацию рецепторного ответа, увеличивает амплитуду последующей гиперполяризации, которая после замены безнатриевой среды на нормальную возвращается к исходному значению. Воздействие ионов Ca2+ на генерацию того же фоторецепторного ответа показано на рисунке 3,Б. Отсутствие этих ионов в омывающем препарат растворе приводило к исчезновению начальной деполяризации и к значительному возрастанию гиперполяризационной фазы реакции. Восстановление исходной реакции достигалось заменой бескальциевого раствора на нормальный.
По аналогичной схеме проводились эксперименты, касающиеся ионной избирательности светозависимых каналов фоторецепторов другого типа, в отличие от первых, не де-поляризующихся, а гиперполяризующихся при освещении. В этих опытах, как и в первом случае, контролировалось влияние ионов Na+ и Ca2+ на состояние фоторецепторного ответа, включающего слабую кратковременную начальную деполяризацию и следующую затем более продолжительную и глубокую гиперполяризацию. Влияние на исследуемый ответ содержания экзогенного Na+ проиллюстрировано на рис. 3,А. Из приведенных записей видно, что выведение Na+ из омывающего клетку раствора, не влияя на начальную деполяризацию рецепторного ответа, увеличивает амплитуду последующей гиперполяризации, которая после замены безнатриевой среды на нормальную возвращается к исходному значению. Воздействие ионов Ca2+ на генерацию того же фоторецепторного ответа показано на рисунке 3,Б. Отсутствие этих ионов в омывающем препарат растворе приводило к исчезновению начальной деполяризации и к значительному возрастанию гиперполяризационной фазы реакции. Восстановление исходной реакции достигалось заменой бескальциевого раствора на нормальный.
Обсуждение
Одним из свойств, отличающих фоторецепторы позвоночных от фоторецепторов беспозвоночных животных, считается полярность их ответа на освещение. Если палочки и колбочки позвоночных животных реагируют на свет гиперполяризаций (см., например: Tomita et al., 1967; Toyoda et al., 1970), то у большинства беспозвоночных освещение вызывает не гипер-, а деполяризацию фоторецепторов. Последнее хорошо изучено на примере Limulus (Fuortes, 1959; Benolken, 1961).
Однако некоторые беспозвоночные демонстрируют исключительные случаи «двойной сетчатки», когда разные электрофизиологические эталоны, характерные для фото-рецепторов беспозвоночных и позвоночных животных, совмещены в одной рецепторной системе. Такой феномен впервые был обнаружен у морского гребешка (Pecten), одна часть фоторецепторов которого реагирует на освещение гиперполяризационным, а другая – де-поляризационным смещением фонового уровня мембранного потенциала. Это было выявлено сначала на интaктной ретине исследуемого моллюска (McReynolds and Gorman, 1970), а в дальнейшем подтверждено и на его изолированных фоторецепторах (Nasi and Gomez, 1992). Противоположным типам электрофизиологических реакций разных фоторецепторов Pecten соответствует разновидность их морфологии. Гиперполяризационный рецепторный ответ обнаруживают фоторецепторы цилиарного типа (характерного для позвоночных), а деполяризационный – классические у беспозвоночных рабдомерные светочувствительные клетки (Musio, 2001).
Сетчатка виноградной улитки (объекта настоящего исследования) включает лишь типично рабдомерные фоторецепторы. Их морфологическое разнообразие сводится к размеру микровилл, одни из которых достигают 100 мнм, а другие несут короткие (до 12 мнм) и нерегулярные микровиллы (Eakin и Brandenburger, 1967). Прямые данные об электрофизиологической неоднородности сетчатки виноградной улитки нами не обнаружены, хотя косвенно об этом свидетельствуют эксперименты H. Gillary (1970). Не вскрывая глазной бокал, H. Gillary впервые зарегистрировал суммарную электрическую активность сетчатки (ЭРГ) этого моллюска, которая меняла свою полярность при перемещении накладного регистрирующего электрода по поверхности глаза. Поскольку сетчатка виноградной улитки однослойная фоторецепторная, данные H. Gillary находят объяснение при допущении, что электрофизиологически неоднородны именно фоторецепторы.
Основным результатом нашей работы является экспериментальное доказательство существования в сетчатке виноградной улитки двух разновидностей клеток, реагирующих на свет противоположно: одни гиперполяризуются, а другие деполяризуются при освещении. Таким образом, сетчатка виноградной улитки Helix pomatia, как и сетчатка гребешка Pecten, может быть отнесена к явлениям «двойной ретины», поскольку включает две группы фоторецепторов с противоположно направленными электрическими реакциями на освещение.
В связи с тем что этот результат получен при исследовании полностью изолированных клеток, гарантированно лишенных синаптических контактов, можно утверждать, что разнонаправленность фоторецепторных ответов (как де-, так и гипер-) у виноградной улитки обусловлена не «сетью» (т. е. межклеточным взаимодействием), а эндогенно, т. е. собственными качествами фоторецепторов. К таким качествам относится и ионная избирательность светочувствительных каналов фоторецепторной мембраны. Одинаковы ли ионы, движение которых создает противоположно направленные фоторецепторные потенциалы виноградной улитки?
В наших экспериментах контролировалось участие ионов натрия и кальция в фоторецепторных ответах обеих противоположных полярностей. Как известно (Бызов, 1971), свет вызывает в фоторецепторах позвоночных животных гиперполяризацию, а в фоторецепторах большинства исследованных беспозвоночных (мечехвост, насекомые и головоногие) – деполяризацию. При этом ионные механизмы генерации рецепторных потенциалов в основном сходны: в обоих случаях свет порождает изменение проводимости натриевых каналов, только у позвоночных они на свету закрываются, а у беспозвоночных – открываются. Известно также, что большая роль в появлении фоторецепторных ответов и в том, и в другом случае принадлежит ионам кальция (Dorlochter and Stieve, 1997; Говардовский, Кузьмин, 1999). Предпринятая в этой работе проверка участия ионов натрия и кальция в генерации как деполяризационных, так и гиперполяризационных ответов фоторецепторов виноградной улитки показала, что в ответ рецепторов, деполяризующихся при освещении, вносят свой вклад как ионы натрия, так и ионы кальция. Этот вывод можно сделать на том основании, что поочередное удаление этих ионов из раствора, омывающего соответствующие фоторецепторы, приводит к падению де-ответа до некоторого остаточного уровня. Далее, поскольку при одновременном исключении Na+ и Ca2+ деполяризационный ответ на освещение блокируется полностью, можно утверждать, что реакция фоторецепторов де-типа целиком обусловлена участием только этих ионов.
Совсем другая картина выявлена при исследовании ионной природы индуцированного светом гиперполяризационного рецепторного ответа. Если раствор вокруг фоторецептора гипер-типа поочередно заменяется на среду, не содержащую Na+ или Ca2+, то и в том, и в другом случае наблюдается не уменьшение (как в случае де-ответа), а увеличение амплитуды реакции. Это увеличение говорит о более сложной ионной природе ответа фоторецепторов гипер-типа, который, вероятно, является суммарным итогом натрий-кальциевой деполяризации и собственно гиперполяризации, вызванной движением каких-то других ионов. Наиболее вероятным кандидатом на роль последних являются ионы калия, точнее фоторецепторная гиперполяризация у виноградной улитки может определяться выходящим калиевым током. Хотя это необычно для позвоночных, у некоторых моллюсков фото-ответ использует калиевые каналы. Одним из таких моллюсков является гребешок, имеющий два вида топически разведенных фоторецепторов – деполяризующихся проксимальных и гиперполяризующихся дистальных. Гиперполяризация в дистальных фото-рецепторах гребешка возникает вследствие увеличения проводимости по калию (Gomez and Nasi, 1995). По-видимому, и гиперполяризующиеся при освещении фоторецепторы виноградной улитки обладают аналогичным механизмом. Таким образом, гипер-ответ фоторецепторов виноградной улитки при дефиците (исключении) Na+ и Са2+ связан с тем, что в норме при освещении фоторецептора ответная калиевая волна гиперполяризации частично компенсируется деполяризацией, вызванной входом ионов натрия или кальция. Это значит, что у фоторецепторов гипер-типа при действии света относительно небольшое число натриевых и кальциевых каналов все же открывается, хотя выход калия при этом доминирует.
Обобщая полученные данные, можно утверждать, что в сетчатке виноградной улитки присутствуют два типа фоторецепторов, характеризующихся разными ионными механизмами генерации рецепторного потенциала и, как следствие, электрической реакцией на освещение противоположной полярности. В результате на следующий уровень зрительной системы от одной популяции фоторецепторов поступает деполяризация, а от другой – гиперполяризация. Фоторецепторные потенциалы разной полярности могут оказывать на структуры следующего уровня, способные к генерации потенциалов действия, два легко различимых эффекта: создание возбуждения одних и подавление активности других элементов. Эти элементы называются соответственно «яркостными» и «темновыми». Подавление спайкового разряда «темнового» (максимально активного в темноте) нервного элемента в период освещения это то, что может достигаться первичной фоторецепторной гиперполяризацией.
Таким образом, разные типы фоторецепторов упрощенной сетчатки виноградной улитки могут являться отправной точкой формирования «яркостной» и «темновой» систем. В зрительных системах других животных, сетчатка которых значительно сложнее, «яркостный» и «темновой» каналы формируются не на рецепторном, а на более высоком, т. е. нейрональном, уровне. Однако общий принцип состоит в том, что кооперация именно этих систем, отражающаяся в двухмерности сферической модели восприятия интенсивности света, создает векторный код различения яркости, который, по-видимому, универсален.
Выводы
1. Ахроматическая зрительная система виноградной улитки содержит два типа фоторецепторов: одни деполяризуются, а другие гиперполяризуются при освещении.
2. Противоположная полярность рецепторных ответов на освещение определяется не межклеточными взаимодействиями, а эндогенными свойствами фоторецепторов виноградной улитки.
3. При формировании рецепторного ответа на мембранах фоторецепторов де- и гипер- типа задействуются разные композиции ионных каналов.