Введение
Формирование и становление ребенка невозможно без взаимодействия с людьми и включения в культурную среду, однако существуют нарушения развития, в том числе расстройства аутистического спектра (РАС), при которых данные процессы нарушается [Кубасова Т.С, 2022]. РАС проявляются уже в раннем детстве [Пахтусова, 2023] и число детей с таким диагнозом постоянно растет [Бронников, 2022]. Для РАС характерны затруднения с регуляцией возбуждения и переключением внимания [Строганова, 2015], снижение внимания к диалогу других людей [Hofsten, 2022] и к подражанию их действиям [Kilroy, 2022], что приводит к выраженному и всестороннему дефициту социального взаимодействия [Божкова, 2020; Духанина, 2023; Морозов С.А, 2023]. Около двух десятилетий назад была сформулирована «гипотеза разбитого зеркала», согласно которой в основе по крайней мере части этих поведенческих особенностей лежат нарушения функционирования т.н. зеркальных нейронов или зеркальной системы мозга (ЗСМ) [Oberman, 2005; Williams, 2001]. Принято считать [Bonini, 2022], что ядро ЗСМ человека состоит из областей неокортекса, расположенных билатерально в нижней теменной доле и вентральной премоторной коре, а в расширенную часть входят сенсомоторная сеть (включающая, помимо указанных областей, верхнюю часть верхней височной извилины, префронтальную, моторную и соматосенсорную кору), а также эмоциональная сеть (включает переднюю поясную кору, миндалевидное тело и островок). Результаты большого числа исследований приводят к выводу, что ЗСМ имеет важное значение для понимания действий и их имитации, формировании гипотетического конструкта «теории сознания» (“Theory of Mind”), способного объяснять и прогнозировать поведение другого [Лебедева, 2017; Kemmerer, 2021], становления социального и просоциального поведения в процессе онтогенеза [Павленко, 2023], т.е. тех сторон психической жизни, которые нарушены при РАС.
Для изучения активности ЗСМ у взрослых нормотипичных испытуемых и людей с РАС в последнее время эффективно применяют различные виды функциональной магнитно-резонансной томографии (ФМРТ) и магнитоэнцефалографию [Chan, 2020; Heyes, 2022]. Однако, при изучении нейрофизиологических механизмов поведения детей дошкольного возраста до сих пор широко используют регистрацию электроэнцефалограммы (ЭЭГ), в частности потому, что данный метод почти не сковывает ребенка, не ограничивает его подвижность. В качестве показателя активации ЗСМ при этом рассматривают десинхронизацию или подавление амплитуды мю-ритма ЭЭГ в альфа-диапазоне частот, поскольку такой феномен выявлен у испытуемых в центральных отведениях при выполнении действия, наблюдении за ним или его мысленном представлении [Ларионова, 2022; Fox, 2016].
Однако до настоящего времени результаты исследований в этой области остаются противоречивыми. Так, в ранних работах отмечена пониженная реактивность ЭЭГ в диапазоне мю-ритма у детей и взрослых с РАС при наблюдении за движениями рук, что расценили как нарушение у них функций ЗСМ [Oberman, 2005]. Однако позже другие исследователи в подобных ситуациях не выявили различий в динамике мю-ритма у взрослых и детей с РАС по сравнению с испытуемыми контрольной группы [Fan, 2010; Sotoodeh, 2019]. Кроме того, в ряде работ, проведенных с применением магнитоэнцефалографии и ФМРТ, у детей и подростков с РАС при выполнении задач, включающих восприятие и имитацию движений, отмечена повышенная, по сравнению с нормотипичными испытуемыми, активация зон неокортекса, являющихся частью ЗСМ [Buard, 2018; Wadsworth, 2018].
Недавний мета-анализ значительного числа исследований с применением ФМРТ в целом подтвердил аномальный характер активации ЗСМ у людей с РАС при наблюдении за действиями окружающих [Chan, 2020]. При этом отмечено, что проявления дисфункции ЗСМ определяются социально-эмоциональным контекстом ситуации. Так наблюдение за действиями, лишенными эмоциональной окраски, сопровождается гиперактивацией областей ЗСМ в левом полушарии (нижняя теменная доля и дополнительная двигательная область), в то время как правая средняя затылочная извилина и левая постцентральная извилина были гипоактивированными по сравнению с активностью неокортекса у типично развивающихся людей. Что касается наблюдения за эмоциональными стимулами, то испытуемые с РАС отличались гиперактивацией правой нижней лобной извилины. Выявленную гиперактивацию участков неокортекса расценивают как свидетельство того, что людям с РАС требуются дополнительные умственные усилия для наблюдения за стимулами и в обычных, и в эмоциональных ситуациях.
Учитывая эти данные, а также многочисленные факты зависимости реакций ЗСМ у нормотипичных людей от характера экспериментальной ситуации, степени вовлеченности в нее и даже от отношения к ее участникам [Kemmerer, 2021], на смену модели «разбитого зеркала», отводящей основную роль в патогенезе аутизма нарушениям функций ядра ЗСМ, приходят другие модели. Согласно им, ядро ЗСМ играет важную роль в низкоуровневой обработке наблюдаемых действий (например, различение конкретных действий), но не в интерпретации действия. Зеркальные нейроны являются частью сложной системы управления. Изменения в поведении у людей с РАС может быть результатом нарушения контроля ядра ЗСМ сверху вниз, со стороны префронтальных и иных областей коры [Heyes, 2022; Khalil, 2018; Yates], входящих в так называемую расширенную ЗСМ. Как следует из этих моделей, активность ЗСМ отражает степень активации и особенности функционирования обширных нейронных сетей неокортекса, вовлеченных в восприятие действий других людей и их имитацию. В последние десятилетия достижения в области социальных наук показали, что имитация является одним из центральных механизмом возникновения уникальных социально-когнитивных способностей человека, и раскрытие онтогенеза имитации имеет важное значение для понимания человеческой культурной эволюции [Essler, 2023].
В связи с этим, особый интерес представляет особенности динамики мю-ритма ЭЭГ детей с РАС в различных социальных или игровых ситуациях, включающих наблюдение за действиями окружающих и их имитацию. При этом нужно учитывать, что именно в ситуации личного социального взаимодействия активация ЗСМ у взрослых и особенно у детей происходит более выражено [Лебедева]. Недавними исследованиями установлено, что при наблюдении за мнимым и реальным действием взрослого экспериментатора, а также при выполнении ребенком самостоятельного действия в игровой ситуации мю-ритм ЭЭГ у нормотипичных детей в возрасте от полутора до трех с половиной лет значимо снижается, причем это снижение было более выражено у детей с высоким развитием рецептивной речи [Михайлова, 2020]. Однако насколько нам известно анализ реактивности мю-ритма ЭЭГ, отражающего степень активации ЗСМ, а также анализ динамики других ритмов ЭЭГ в описанных трех ситуациях у детей дошкольного возраста с РАС ранее не проводился.
В связи с этим целью исследования является анализ динамики ритмов ЭЭГ у нормотипичных детей и детей с РАС при наблюдении за мнимым действием, реальным действием и повторении действия.
Организация и методы исследования
Исследование было проведено на базе Центра коллективного пользования научным оборудованием «Экспериментальная физиология и биофизика» Крымского федерального университета имени В. И. Вернадского. Основную группу детей с диагнозом РАС в возрасте 4–7 лет составили 20 детей (из них 14 детей мужского и 6 – женского пола). Средний возраст детей с РАС – 71,7 ± 13,3 месяцев. Группу сравнения составили 33 типично развивающихся сверстника (из них 18 мальчиков и 15 девочек). Средний возраст детей данной группы составил 71,3 ± 13,9 месяцев.Родители детей дали письменное информированное согласие на участие ребенка в экспериментах. Настоящее исследование соответствовало этическим принципам Хельсинкской декларации 1964 г. и было одобрено этическим комитетом Крымского федерального университета им. В.И. Вернадского.Критерии включения детей в группу сравнения: достаточный уровень когнитивного развития (IQ от 90 до 120 баллов по тесту Векслера в вариантах WISC и WPPSI); отсутствие хронических заболеваний нервной системы. Критерии включения для детей основной группы: наличие установленного детским психиатром диагноза РАС, подтвержденного методиками ADOS-2 (балл от 3 до 10) и CARS (баллы от 30 до 60). Критерии включения для детей обеих групп: предпочтение правой руки; наличие нормального уровня зрения и слуха.Во время проведения дети 4-7 лет сидели на стуле перед столом, на котором располагался стимульный материал. Исходная (фоновая) ЭЭГ была зарегистрирована при просмотре видеозаписи вращающегося мяча в состоянии устойчивого зрительного внимания (УЗВ) ребенка. При этом ребенку давали инструкцию «посиди спокойно, глядя на мячик».
Затем проводили регистрацию ЭЭГ в трех экспериментальных ситуациях по ранее предложенной методике [Михайлова, 2020; Nyström, 2011]:
- «Наблюдение мнимого действия» – ребенок наблюдает, как экспериментатор проводит рукой по установленной на столе горке (3 повторения). Считается, что ребенок не видит в этом действии реальной цели [Nyström, 2011].
- «Наблюдение реального действия» – ребенок наблюдает, как экспериментатор сталкивает с горки игрушечную машинку (3 повторения).
- «Выполнение действия» – экспериментатор ставит горку с машинкой перед ребенком, и ребенок сам ее сталкивает (3 повторения).
Перед ситуациями 1 и 2 давалась инструкция: «смотри внимательно», а перед ситуацией 3: «сделай, как делал я». Понимание инструкций подтверждалось тем, что все дети выполняли соответствующее задание.
ЭЭГ была зарегистрирована с помощью электроэнцефалографа «Нейрон-Спектр-3» («Нейрософт»; Россия) монополярно в лобных, центральных, височных, теменных и затылочных отведениях в соответствии с международной системой «10-20». Полоса пропускания сигнала составила 0,5–30,0 Гц при частоте дискретизации 250 Гц. В рамках настоящего исследования проводили анализ ЭЭГ в следующих отведениях: F3, Fz, F4, F7, F8, C3, Cz, C4, P3, Pz, P4. Указанные регионы выбраны как области интереса на основании данных литературы о целесообразности анализа мю-ритма у детей не только в центральных, но и во фронтальных и теменных областях [Marshall, 2011; Warreyn, 2013]. В качестве референтного использовался объединенный ушной электрод. Во время записи ЭЭГ велась синхронная видео- и аудиорегистрация. Фрагменты ЭЭГ с большим количеством артефактов, а также пробы, в которых видеозапись показала отсутствие внимания ребенка к ситуации, были исключены из анализа.
Для анализа исходной ЭЭГ использовался отрезок записи длительностью 20-30 с. Также анализировались отрезки ЭЭГ длительностью 20-30 с в трех описанных выше экспериментальных условиях. Начало фрагмента было положено за 1 с до момента первого прикосновения руки экспериментатора к горке или к машинке, а также прикосновения ребенка к машинке. Завершение отрезка – через 1 с после последнего прикосновения руки экспериментатора к горке или к машинке, а также прикосновения ребенка к машинке. Указанные фрагменты ЭЭГ подвергались быстрому преобразованию Фурье с эпохой анализа 2 с и взаимным перекрытием эпох 50%. Обработку данных ЭЭГ проводили с помощью программы WinEEG («Мицар»; Россия).
Индивидуальный для каждого отдельного испытуемого диапазон мю-ритма определяли следующим образом. В условиях УЗВ на спектре ЭЭГ в отведении С3 выявляли пик мощности в диапазоне 6-14 Гц, который исчезал во время совершения ребенком движения рукой. На усредненный по последовательным эпохам спектр ЭЭГ, зарегистрированной в ситуациях УЗВ, накладывали усредненный по трем повторениям спектр ЭЭГ, зарегистрированной в условиях «Выполнения действия». За границы анализируемого диапазона принимали точки пересечения спектров. Данную компоненту альфа-ритма, демонстрирующую супрессию мощности в ситуации двигательной активности, рассматривают как сенсомоторный или мю-ритм [Thorpe, 2016]. Для индивидуально-определенной частотной полосы мю-ритма в программе WinEEG («Мицар»; Россия) была рассчитана амплитуда, которая затем была прологарифмирована для нормализации распределения (lg мкВ).
Для оценки реакций десинхронизации у детей исследованных групп рассчитывали индексы реактивности (ИР) мю-ритма в соответствии с формулой [k = ln(B/A)], где k – коэффициент реактивности мю-ритма, B – мощность мю-ритма в экспериментальной ситуации, A – мощность мю-ритма в фоновой ситуации, подобно тому, как это делали при анализе ЭЭГ-реакций, сопровождающих просоциальное поведение [Павленко, 2023а]. Для экспериментальных условий «Наблюдение мнимого действия», «Наблюдение реального действия», «Выполнение действия» индексы реактивности обозначали как ИР-1, ИР-2, ИР-3, соответственно.
Значения ИР > 0 свидетельствуют об увеличении мощности мю-ритма в экспериментальной ситуации по сравнению с фоновым уровнем (синхронизация), а ИР < 0 – о снижении мощности мю-ритма (десинхронизация).
Статистическая обработка данных
Статистическая обработка данных была произведена в программе STATISTICA 12.0 (StatSoft Inc.; США). Результаты измерений были представлены как средние значения ± стандартная ошибка среднего. Изменения амплитуды мю-ритма были проанализированы во всех экспериментальных ситуациях относительно друг друга и по отношению к УЗВ с помощью дисперсионного анализа ANOVA с повторными измерениями (repeated measures) с факторами СИТУАЦИЯ (2 уровня), ЛОКУС (уровни: 11 отведений) и СИТУАЦИЯ × ЛОКУС для каждой группы отдельно. С целью оценки эффектов изменений мощности мю-ритма в каждом из 11 локусов ЭЭГ был использован метод априорных контрастов (оценка F-распределения). Для оценки особенностей ИР у детей с РАС определяли влияние межсубъектного фактора ГРУППА (два уровня: дети с РАС и группа сравнения) и внутрисубъектного фактора ЛОКУС (11 отведений ЭЭГ) по схеме 2 × 11. Для расчета статистической значимости различий ИР в привязке к каждому из 11 отведений ЭЭГ использовали метод линейных контрастов. Статистически значимыми считали различия при р < 0,05 и p< 0,01.
Результаты исследования
Были определены показатели индивидуально определенного мю-ритма у детей исследуемых групп. Среднее значение пиковой частоты индивидуально определенного мю-ритма для нормотипичных детей составило 9,3±0,2, а для детей с РАС – 10,0 ±0,3 Гц. Распределение показателей не отличалось от нормального (критерий Шапиро-Уилки). Применение Т-критерий Стьюдента значимых различий пиковой частоты мю-ритма у детей двух групп не выявило. Среднее значение пиковой частоты индивидуально определенного мю-ритма для всей совокупности испытуемых составило 9,5±0,2, нижней границы частотного диапазона мю-ритма – 8,1±0,2, верхней границы – 11,1±0,2 Гц.Дисперсионный анализ repeated measures ANOVA позволил выявить влияние факторов СИТУАЦИЯ, ЛОКУС и их взаимодействия на показатели амплитуды мю-ритма ЭЭГ. В табл. 1 представлены результаты анализа изменений амплитуды ЭЭГ в частотном диапазоне мю-ритма в трёх упомянутых выше ситуациях: «Наблюдение мнимого действия», «Наблюдение реального действия», «Выполнение действия» относительно ситуации УЗВ с учетом локуса отведения для группы нормотипичных детей. На амплитуду ЭЭГ в диапазоне мю-ритма значимо влияют факторы СИТУАЦИЯ и ЛОКУС, а также их взаимодействие. Как показано на рис. 1 у детей контрольной группы амплитуда ЭЭГ в диапазоне мю-ритма статистически значимо снижается в трех исследуемых экспериментальных ситуациях во всех отведениях, за исключением локусов F7 и F8 (метод априорных контрастов).
Таблица 1. Результаты дисперсионного анализа изменений мощности ЭЭГ в диапазоне мю-ритма в ситуациях «Наблюдение мнимого действия», «Наблюдение реального действия» и «Выполнение действия» относительно исходной записи в условиях устойчивого зрительного внимания (УЗВ) у типично развивающихся детей
|
Сравниваемые ситуации |
СИТУАЦИЯ |
ЛОКУС |
СИТУАЦИЯ × ЛОКУС |
|||
|
F1.32 |
р |
F10.320 |
p |
F10.320 |
P |
|
|
«Наблюдение мнимого действия» и УЗВ |
10,77 |
0,003 |
18,72 |
< 0,001 |
6,05 |
< 0,001 |
|
«Наблюдение реального действия» и УЗВ |
23,15 |
< 0,001 |
18,11 |
<0,001 |
6,89 |
< 0,001 |
|
«Выполнение действия» и УЗВ |
23,19 |
< 0,001 |
18,36 |
< 0,001 |
12,27 |
< 0,001 |
В табл. 2 представлены результаты анализа repeated measures ANOVA изменений амплитуды ЭЭГ в частотном диапазоне мю-ритма в трёх описанных выше ситуациях для группы детей с РАС. Для всех экспериментальных ситуаций выявлено значимое влияние фактора СИТУАЦИЯ и ЛОКУС. Метод априорных контрастов показал, что у детей с РАС амплитуда ЭЭГ в диапазоне мю-ритма во всех экспериментальных ситуациях значимо снижается в большинстве локусов (рис. 2). Влияния взаимодействия факторов СИТУАЦИЯ × ЛОКУС во всех сравниваемых ситуациях не имели значимого эффекта.
Таблица 2. Результаты дисперсионного анализа изменений мощности ЭЭГ в диапазоне мю-ритма в ситуациях «Наблюдение мнимого действия», «Наблюдение реального действия» и «Выполнение действия» относительно исходной записи в условиях устойчивого зрительного внимания (УЗВ) для детей с РАС
|
Сравниваемые ситуации |
СИТУАЦИЯ |
ЛОКУС |
СИТУАЦИЯ × ЛОКУС |
|||
|
F1.18 |
p |
F10.150 |
p |
F10.150 |
P |
|
|
«Наблюдение мнимого действия» и УЗВ |
9,72 |
0,006 |
24,2 |
< 0,001 |
0,41 |
0,94 |
|
«Наблюдение реального действия» и УЗВ |
5,53 |
0,03 |
27,52 |
< 0,001 |
0,69 |
0,74 |
|
«Выполнение действия» и УЗВ |
7 |
0,02 |
21,71 |
< 0,001 |
0,81 |
0,62 |
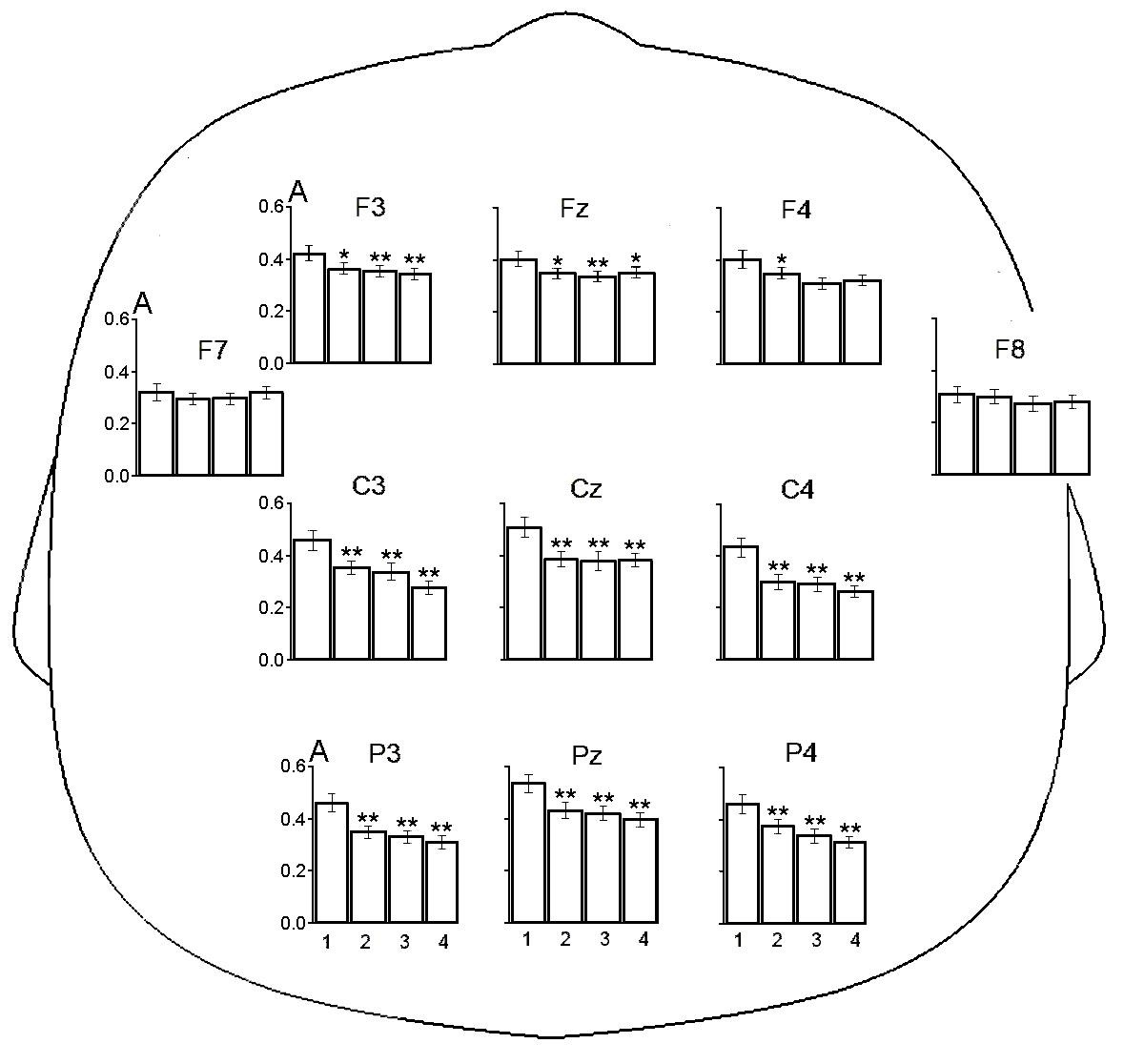
Рис. 1. Амплитуда ЭЭГ (А, lg мкВ) в диапазоне мю-ритма в ситуациях устойчивого зрительного внимания (1), наблюдения мнимого действия (2), наблюдения реального действия (3) и выполнения самостоятельного действия (4) у типично развивающихся детей. Представлены средние значения ± стандартная ошибка среднего. Звездочками отмечены случаи достоверных различий между исходной ситуацией (1) и ситуацией наблюдения и выполнения движений (2,3,4), выявленные методом априорных контрастов (* – р < 0.05, ** – р < 0.01)
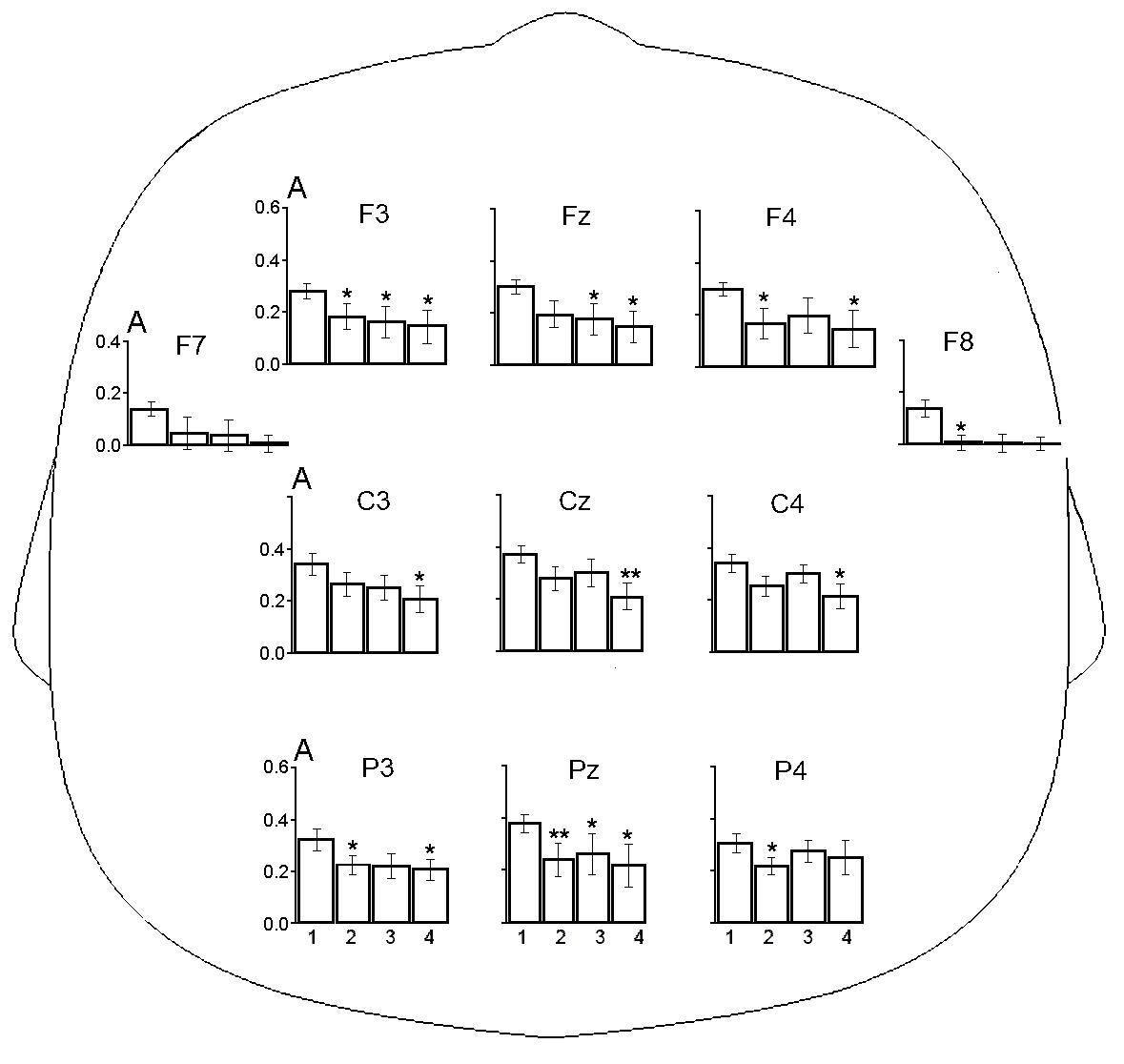
Рис. 2. Амплитуда ЭЭГ (А, lg мкВ) в диапазоне мю-ритма в ситуациях устойчивого зрительного внимания (1), наблюдения мнимого действия (2), наблюдения реального действия (3) и выполнения самостоятельного действия (4) у детей с РАС. Остальные обозначения как на рис. 1
Для сравнения изменений мю-ритма ЭЭГ у исследуемых группа детей в ситуациях «Наблюдение мнимого действия», «Наблюдение реального действия» и «Выполнение действия» был проведен дисперсионный анализ ИР с учетом факторов ГРУППА (нормотипичные дети и дети с РАС), ЛОКУС и их взаимодействия (табл. 3). Выявлено значимое влияние взаимодействия факторов ГРУППА × ЛОКУС на ИР-2 и ИР-3, а также значимое влияние фактора ЛОКУС на ИР-3.
На рис. 3 представлены диаграммы ИР ЭЭГ в диапазоне мю-ритма в ситуациях «Наблюдение мнимого действия», «Наблюдение реального действия», «Выполнение действия» относительно УЗВ у детей исследуемых групп. Дальнейший анализ методом линейных контрастов выявил значимые различия в показателях ИР-2 в отведении С4. Как видно из рисунка 3 в данном локусе у детей с РАС обнаружена меньшая степень десинхронизации мю-ритма по сравнению с группой сравнения в ситуации «Наблюдение реального действия».
Таблица 3. Результаты дисперсионного анализа индексов реактивности ЭЭГ в диапазоне мю-ритма в зависимости от факторов ГРУППА (нормотипичные дети и дети с РАС) и ЛОКУС (11 отведений ЭЭГ)
|
Индексы реактивности |
ГРУППА |
ЛОКУС |
ГРУППА × ЛОКУС |
|||
|
F1.49 |
p |
F10.480 |
P |
F10.480 |
P |
|
|
ИР-1 |
0,004 |
0,95 |
1,17 |
0,31 |
2,19 |
0,02 |
|
ИР-2 |
0,07 |
0,79 |
1,08 |
0,37 |
3,42 |
<0,001 |
|
ИР-3 |
0,25 |
0,62 |
3,04 |
<0,001 |
3,85 |
<0,001 |
Примечание: ИР-1, ИР-2, ИР-3 – индексы реактивности для ситуаций «Наблюдение мнимого действия», «Наблюдение реального действия», «Выполнение действия», соответственно.
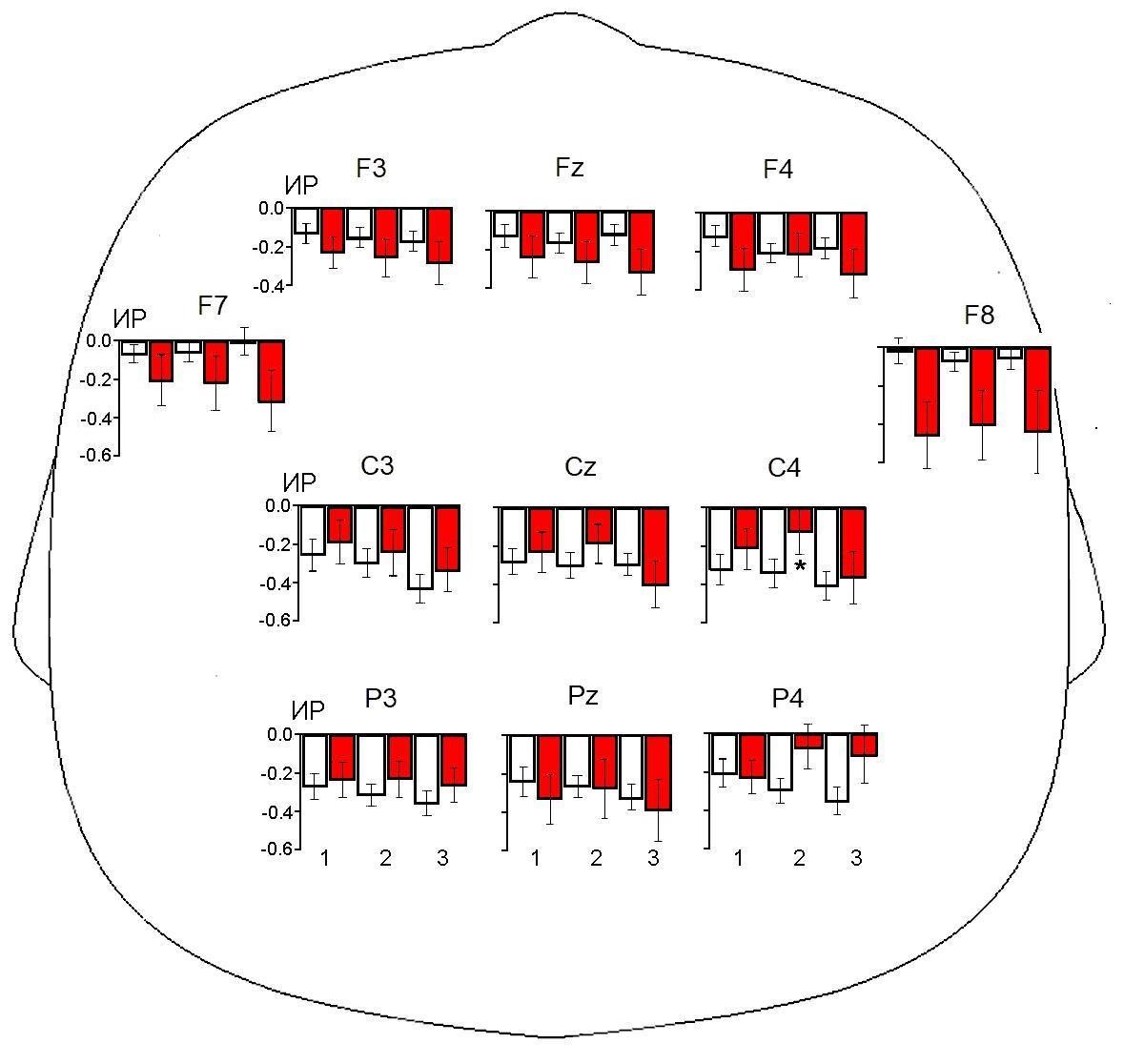
Рис. 3. Индексы реактивности ЭЭГ в диапазоне мю-ритма в группах нормотипичных детей (белые столбцы) и детей с РАС (красные столбцы) в условиях наблюдения мнимого действия (1), наблюдения реального действия (2) и собственного выполнения действия (3) относительно ситуации устойчивого зрительного внимания. Остальные обозначения как на рис. 1
Обсуждение результатов исследования
У типично развивающихся детей дошкольного возраста и у их сверстников с РАС в ситуациях наблюдения за мнимым и реальным действием, а также при выполнении ребенком самостоятельного действия было выявлено значимое падение амплитуды мю-ритма ЭЭГ относительно базового уровня, определенного в ситуации УЗВ. Выявленная десинхронизация мю-ритма при выполнении действия и наблюдения за действием у нормотипичных детей соответствует результатам работ других авторов [Cannon, 2016; Filippi, 2016; Marshall, 2011; Saby, 2013]. Обнаруженное снижение амплитуды мю-ритма в указанных ситуациях у детей с РАС также согласуется с наблюдениями ряда исследователей [Fan, 2010; Sotoodeh, 2019]. Поскольку десинхронизацию мю-ритма рассматривают как отражение активации ЗСМ [Ларионова, 2022; Fox, 2016], можно заключить, что в наших экспериментальных ситуациях, которые включали игровой компонент и, предположительно, сопровождались эмоциональным вовлечением испытуемых, ЗСМ не только нормотипичных детей, но и детей с РАС оказалась способна к активации. Однако паттерн ЭЭГ-реакций двух групп детей существенно отличался.
Прежде всего надо отметить у детей с РАС большее снижение амплитуды ЭЭГ в частотном диапазоне мю-ритма в отведении F7 и, особенно, F8 во всех анализируемых ситуациях по сравнению с детьми группы сравнения (см. рис. 1 и 2). Такая десинхронизация данного ритма указывает на повышенную активацию нейронных сетей. Отведении F7 и, F8 расположены над нижними лобными извилинами левого и правого полушарий [Koessler, 2009]. Как уже отмечалось, применение ФМРТ ранее выявило у испытуемых с РАС гиперактивацию правой нижней лобной извилины при наблюдении за эмоциональными стимулами. Вслед за другими авторами [Chan, 2020], выявленную гиперактивацию участков лобной коры можно расценить как свидетельство того, что детям с РАС требуется вовлечение дополнительных нервных ресурсов для восприятия стимулов в эмоциональных ситуациях.
В то же время, в условиях наблюдения за реальным действием у нормотипичных детей по сравнению с группой РАС отмечается значимо большая десинхронизация ЭЭГ в диапазоне мю-ритма в центральных отведениях. Различия достигают статистически значимого уровня в отведении С4 правого полушария (см. рис 3). Данное отведение расположено над постцентральной извилиной [Koessler, 2009]. В этой области неокортекса, как предполагают [Bonini, 2022], находится часть нейронных сетей ЗСМ, отвечающих за представление собственных и чужих действий. Большая реактивность мю-ритма у детей группы сравнения при наблюдении за реальным действием отражает, вероятно, большую активацию ЗСМ, которая позволяет им в реальных жизненных условиях лучше распознавать намерения окружающих. Меньшая десинхронизация мю-ритма в центральных отведениях неокортекса у группы РАС в указанной экспериментальной ситуации может свидетельствовать о менее выраженной реакции ЗСМ на целенаправленные действия других людей.
Нельзя также исключить и другого объяснения меньшего подавления мю-ритма у детей с РАС при наблюдении за реальным действием. Недавно была выдвинута гипотеза о том, что мю-ритм отражает работу воротного механизма передачи информации в развивающейся нервной системе ребенка [Köster, 2023]. Эта гипотеза предполагает, что если снижение мощности мю-ритма указывает на облегчение двигательных процессов (например, при выполнении действия), то увеличение мощности мю-ритма свидетельствует, что двигательные процессы заторможены (например, для предотвращения имитации наблюдаемого действия). Таким образом, динамика амплитуды мю-ритма отражает баланс активации и торможения процессов, лежащих в основе запуска или остановки физических действий. Можно предположить, что детям с РАС, которые наблюдали за действиями взрослого в первых двух экспериментальных ситуациях, было сложнее удерживаться, чем нормотипичным детям, от того, чтобы только наблюдать, а не столкнуть машинку самостоятельно.
Выявленные особенности реактивности ЭЭГ у детей с РАС важны для понимания нарушений социального поведения у детей с данными расстройствами развития, а также могут быть использованы в коррекционной работе, в том числе, при проведении сеансов биологической обратной связи по ЭЭГ.
Выводы
- У нормотипичных детей дошкольного возраста и детей с РАС выявлена десинхронизация мощности ЭЭГ в диапазоне мю-ритма в центральных, фронтальных и теменных отведениях в условиях наблюдения мнимого действия, реального действия и при выполнении ребенком самостоятельного действия. Это указывает на активацию ЗСМ при наблюдении за действиями окружающих как у группы нормы, так и у детей с РАС. При этом паттерн ЭЭГ-реакций двух групп детей существенно отличался.
- У детей с РАС в трех экспериментальных ситуациях наблюдалась бо́льшая, по сравнению с нормотипичными детьми десинхронизация мю-ритма в областях нижних лобных извилин левого и правого полушарий. Выявленная гиперактивация участков лобной коры указывает на необходимость вовлечения дополнительных нервных ресурсов при восприятии стимулов в эмоциональных ситуациях.
- В условиях наблюдения реального действия у детей с РАС в правом центральном отведении отмечается значимо меньшая десинхронизация мю-ритма по сравнению с группой типично развивающихся детей.