Примерно с середины XIX в. основным источником сведений об эволюции жизни на Земле были палеонтология и эмбриология. Сохранившиеся в осадочных пластах Земли фоссилизованные остатки живых существ («окаменелости») и следов их жизнедеятельности, а также повторение, до некоторой степени, в эмбриональном морфогенезе, онтогенезе в целом, эволюционной истории организмов («рекапитуляции») позволили построить развернутую картину эволюции живых существ. Наложение на эту картину результатов исследования поведения и психики существ, как эволюционно близких к человеку (приматы), так и отстоящих далеко, позволило в очень общих чертах описать эволюцию психики человека и некоторых современных ему животных, а также реконструировать отдельные стороны поведения и психики вымерших организмов и на этой основе воссоздать некоторые аспекты эволюционной истории их современных потомков (см. например: Северцов, 1922/1984; Бернштейн, 1947; Hinde, 1966).
Появление новых методов в молекулярной биологии и генетике открыло широкие возможности для исследований, предметом которых стали эволюция «репликаторов»: прежде всего, нуклеиновых кислот, а также эволюция отдельных белков, в перспективе – гормонов, нейротрансмиттеров, нейромедиаторов и других регуляторов и модуляторов активности организмов. В свою очередь, эволюция регуляторных и других систем (например, коммуникативных, когнитивных и др.) тесно связана с эволюцией морфофункциональной организации живых систем.
В своем подходе к эволюционной проблематике мы исходим из представления о психике как свойстве всех без исключения живых систем и об эволюции как о наследуемых изменениях в организации и поведении живых систем, закрепляемых не только генетически, но и эпигенетически, поведенчески, социально и рядом других способов, между собой взаимодействующих, переплетающихся и взаимосвязанных.
Мы исходим также из представления о двух основных линиях организации живых существ и их сообществ: социализации и индивидуализации. В истории жизни на Земле эти две линии тесно переплетены: одноклеточные организмы формируют все более тесно связанные сообщества и многоклеточные организмы, а многоклеточные сегодня представлены как относительно независимыми от конспецификов индивидами, так и более или менее тесно связанными социальными живыми системами – от «рыхлых» субсоциальных до эусоциальных, в которых отдельные особи, будучи изъяты из социального суперорганизма, практически не выживают.
В современной науке накоплены знания об особенностях социальной жизни существ различного уровня развития – насекомых, птиц, рыб, позвоночных животных, людей (Мантейфель, 1980; Панов, 2011; Шовен, 2012; Эспинас, 2012). Обеспечивая целостность и самовоспроизведение живых существ, социальная жизнь необходима для выживания в изменяющемся мире и поэтому обусловлена биологическими механизмами. Все чаще интересы исследователей обращаются к глубоким корням социального поведения, которое характерно и для древнейших существ Земли – цианобактерий (Олескин, 2009; Сумина, 2006). Бактериальные клетки живут почти исключительно в сообществах с другими микроорганизмами, часто в ассоциации с многоклеточными «хозяевами» и не могут существовать в виде изолированных единиц. Многие микроорганизмы создают биопленки, исследования показывают их структурную и физиологическую сложность и разнообразие (биопленки – это плотно упакованные сообщества микробных клеток, которые растут на поверхности и окружают себя секретируемыми полимерами, см. Рыбальченко, 2003). Они действуют подобно многоклеточным организмам, использующим сложные системы межклеточной коммуникации для лучшей приспособленности к меняющимся условиям окружающей среды.
В ходе эволюции организмы выработали способность существовать в относительной независимости от общественной среды. Мы предполагаем, что биологической предпосылкой способа выживания (зависимого или относительно независимого от сообщества) является разнообразие осцилляторных процессов. Эта идея основывается на фактах, полученных в опытах на живых существах разной степени сложности – именно этот вид электрической активности обнаружен у древнейших микроорганизмов Земли – цианобактерий, эндогенные колебания являются первыми электрическими сигналами дифференцирующихся зигот (Кузень и др., 1980), он идентифицирован у нейронов различных отделов нервной системы многоклеточных существ и выполняет разнообразные функции (Греченко и др., 2014; Данилова и др., 2013). Эндогенные электрические осцилляторы являются результатом внутриклеточной химической сигнализации, обеспечивающей жизнедеятельность организма (Веселкин, Наточин, 2010). Эволюция химической сигнализации шла по пути развития механизмов ее регуляции – чем сложнее информационные процессы, происходящие во внутренней среде клетки живого организма, тем шире частотный репертуар его осцилляторных систем и разнообразней паттерны их проявления, тем выше его шансы на независимое существование. От наличия эндогенных осцилляторов и их свойств зависит разнообразие и реализация поведенческих программ индивидуальных организмов (Соколов, 1974).
Для проверки гипотезы в экспериментах регистрировали электрическую активность от индивидуальных организмов и от их скопления (колонии или сообщества) у эволюционно древних существ: прокариот – цианобактерий Oscillatoria terebriformis и Geitlerinema sp. и Halothece sp., одноклеточных эукариот – дрожжей Saccharomyces cerevisiae и инфузорий Paramecium caudatum, а также у моллюсков Helix pomatia и Helix lucorum.
Метод
Применялся метод регистрации электрической активности при помощи стеклянных микроэлектродов. Для работы с цианобактериями Oscillatoria terebriformis применяли физраствор (в граммах на литр): NaHCO3 – 3, Na2CO3 – 17, K2HPO4 – 0,5, NaCl – 30, KNO3 – 2,5, MgSO4 – 0,2, CaCl2 – 0,04, FeSO4 – 0,01. Фрагмент цианобактериального мата, сформированного нитчатыми цианобактериями Geitlerinema sp. и содержавшего одиночные цианобактерии Halothece sp. и одноклеточные водоросли Niscia, исследовался в пробе воды из естественной среды обитания (оз. Дусь-Холь, 45 км. южнее г. Кызыл, Республика Тыва), представляющую собой естественный рассол – хлоридно-магниево-натриевую рапу с минерализацией до 125 граммов на литр и более; анионы Cl (62-73), SO (27-38); катионы Na+K (70-76), Mg (24-29); pH раствора 7,9. Электроды заполняли 1 М раствором KCl.
В опытах на дрожжах Saccharomyces cerevisiae использовалась культура «диких» дрожжей, помещенных в водную среду температурой 22–25о С. Регистрация электрической активности производилась МЭ, заполненными 1 М KCl.
Электрическая активность свободноживущих инфузорий Paramecium caudatum регистрировалась в камере объемом 0,35 мл. При помощи микропипетки парамеций переносили в экспериментальную камеру, содержавшую специальный физиологический раствор (KCl – 4 mM, CaCl – 1 mM, MgCl – 5 mM, tris HCl – 1 mM. pH раствора – 7,2). Для регистрации применяли стеклянные микроэлектроды (МЭ), заполненные 0,1 М КСl. Для электрической стимуляции использовали деполяризационные импульсы тока силой 0,1–1 нА, длительностью 1000–3000 мс.
Для работы на нервной системе и полностью изолированных клетках моллюска Helix lucorum применяли стандартную технику микроэлектродной регистрации электриче- ской активности нейронов. Методика подробно описана в ранее опубликованных работах (Греченко, Соколов, 1979).
Результаты
Пейсмекеры прокариот. Цианобактерии Oscillatoria terebriformis, Geitlerinema sp. и Halothece sp. – фотосинтезирующие прокариоты, эволюционный возраст которых составляет около 3,5 млрд лет. В природе образуют пленки и цианобактериальные маты.
Цианобактерии могут приспосабливаться к широкому ряду изменений окружающей среды за счет большой адаптивности поведения и своего рода «социальных» отношений в формируемых ими сообществах (Сумина, 2006).
Электрофизиологические опыты позволили обнаружить информационные обмены между членами сообщества. При внутриклеточной регистрации электрической активности от индивидуальной цианобактериальной нити обнаружен стабильный уровень мембранного потенциала (МП) и частоты осцилляций, которые сохранялись в течение более 2 часов. Частота осцилляций – 0,1–0,5 Гц и амплитуда – до 50 мВ (рис. 1, б). Регистрация электрической активности макроэлектродом в цианобактериальной пленке показывает синхронизированные синусоидальные ритмы частотой от 3 Гц до 35–40 Гц. Когда деятельность членов цианобактериального сообщества направлена на решение жизненно важной задачи (например, освоение новых поверхностей, противостояние другому сообществу), их актив- ность синхронизирована, что выражается в генерации высокоамплитудных ритмических колебаний суммарного потенциала (рис. 1, а).
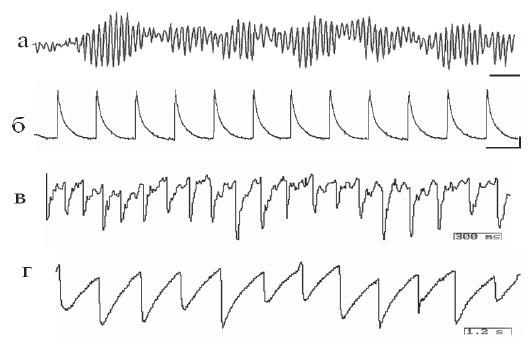
Рис. 1. Электрическая активность цианобактерий Oscillatoria terebriformis – суммарная (а) и внутриклеточная (б) и цианобактерий Geitlerinema sp. и Halothece sp., входивших в состав пробы цианобактериального мата, – суммарная (в) и внутриклеточная (г). Калибровка: а, б, в – 300 мс, г – 1,2 с; б – 10 мВ
За счет движения нитей и деления клеток в биопленке создаются органоподобные образования. При регистрации потенциалов в зонах, заведомо отличающихся активностью элементов, например, в области формирования тяжей (структурных элементов) и в зонах спокойствия, были получены разные по уровню синхронизированной активности явления. Чем сложнее внутренняя организация живого существа, тем в большей степени метаболические процессы отражаются в эндогенной ритмике: они актуализируются в электрической активности клеток и включают определенные системы исполнения. В наших экспериментах показано, что цианобактерии имеют чрезвычайно устойчивую внутриклеточную электрическую активность, характеризующуюся постоянством периодичности (рис. 1, а, в). Синхронизированная электрическая активность является пусковым механизмом совместной деятельности. Как показывают исследования, бактериальные биосоциальные системы характеризуются единым жизненным циклом, следовательно, их активность упорядочить во времени проще, чем, например, у дрожжей. У эволюционно более продвинутых дрожжевых клеток внутриклеточная регистрация выявляет наличие сложных паттернов эндогенной активности (рис. 2, а, б), среди которых сложно выделить электрические процессы, выражающие циркадианные ритмы.
Пейсмекеры дрожжей (одноклеточные эукариоты, грибы). Относительно времени появления организмов этой группы в геологической летописи Земли достоверных данных нет: вероятно, это произошло в интервале от 2 млрд до 650 млн лет назад. Размеры дрожжевых клеток обычно составляют 3–7 мкм в диаметре. Мембрана дрожжевых клеток трудно проницаема для микроэлектрода, так как содержит хитин. Осцилляции, зарегистрированные внутриклеточным микроэлектродом от индивидуальной дрожжевой клетки, характеризуются чрезвычайным разнообразием как по амплитуде, так и по форме и временной организации. На электрограммах четко различима электрическая активность осцилляторов, имеющая сложные паттерны (рис. 2, а).
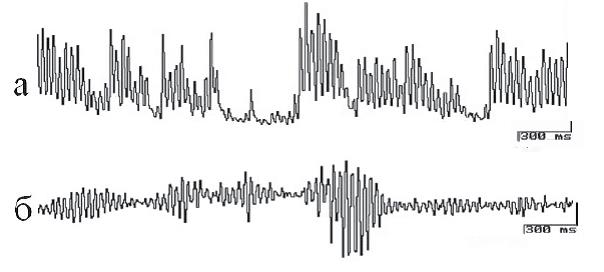
Рис. 2. Электрическая активность дрожжевых клеток Saccharomyces cerevisiae, отведенная внутриклеточным микроэлектродом (а) и макроэлектродом (б). Калибровка: 300 мс, 10 мВ (а), 50 мВ (б)
Паттерны активности повторяются с определенной частотой – каждый вид отличаетcя индивидуальной периодичностью. На электрические процессы дрожжевых клеток влияют такие факторы внешней среды, как температура, состав жидкостной среды в экспериментальной камере и длительность нахождения в растворе определенного состава. Регистрация суммарной активности обнаруживает у дрожжей высокочастотный компонент до 30 Гц и низкочастотные компоненты около 0,1 Гц (существуют ритмические компоненты с частотой 1/час). В ряде случаев генерация высокочастотных колебаний развивается упорядоченно, группы следуют с частотой 0,8–0,9 Гц (рис. 2). В результате проведенных опытов показано, что величина заряда клетки изменяется в зависимости от цикла активности, осуществляемого организмом.
Пейсмекеры парамеций. Это свободноживущие одноклеточные эукариоты, наиболее древние представители которых появились примерно 2,5 млрд лет назад. Клетки имеют форму туфельки длиной 0,12–0,13 мм. Исследователями накоплен громадный материал о структуре различных клеточных элементов, свойствах электровозбудимой цитоплазматической мембраны, белковом составе и свойствах отдельных белков, входящих в систему управления двигательной активностью парамеций. Микроэлектродные регистрации, предпринятые для исследования электрических процессов, происходящих в единственной клетке этих живых существ, позволили обнаружить взаимодействие множества эндогенных осцилляторов. Функция каждого из осцилляторов, по-видимому, связана с контролированием движений определенной группы ресничек парамеции. Частота зарегистрированных колебаний – от 1/с до 50–55/с, амплитуда – от 2–3 мВ до 30 мВ (рис. 1, б). Результаты регистрации электрической активности Paramecium caudatum внутриклеточным микроэлектродом показали, что уровень мембранного потенциала покоя (ПП) этих существ характеризуется значительными колебаниями: обнаружены изменения от –40–45 мВ до +20 мВ, периоды стабильного значения ПП чрезвычайно короткие – в условиях проведенных опытов не более 30–70 с. Регистрируются медленные и быстрые осцилляции, как правило, одновременно активны несколько эндогенных осцилляторов. Эти данные подтверждаются и другими исследователями, экспериментировавшими на парамециях (Доронин, Зазулин, 1976). Одновременно работает два–три осциллятора, каждый из них характеризуется своей частотой, формой и ампли- тудой (рис. 3, а, б). Увеличение амплитуды волн эндогенной активности может приводить к генерации потенциалов действия (ПД), отличающихся амплитудой, которая, по-видимому, зависит от функциональных особенностей вызвавшего его пейсмекера.
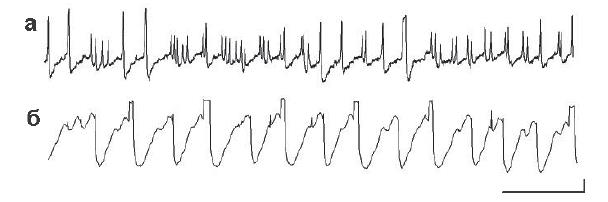
Рис. 3. Электрическая активность, зарегистрированная от инфузорий Paramecium caudatum внутриклеточным микроэлектродом. Показаны различные паттерны активности, отведенные от одного и того же микроорганизма в разное время опыта. Калибровка: 200 мс, 10 мВ
На электрограммах можно видеть ПД как одинаковой, так и различной амплитуды, отличающиеся единицами и даже десятками мВ (рис. 3). Частота зарегистрированных колебаний – от 0,1 Гц до 20 Гц, амплитуда – от 2–3 мВ до 50 мВ. В ряде работ показано, что движения ресничек у парамеции, расположенных в разных частях тела, различаются частотой биений более чем в два раза (Eckert, Naitoh, 1970). Анализ экспериментальных данных, полученных при изучении поведения свободно плавающих одноклеточных, позволяет сделать вывод о том, что система управления движениями должна быть многоуровневой и включать в себя рецепторное, афферентное центральное, эфферентное и эффекторное звенья. При этом роль центрального интегратора и координатора двигательного поведения может выполнять ядро клетки, тесно связанное с периферией с помощью динамичных элементов цитоскелета (Свидерский и др., 2007). Действие деполяризационного тока синхронизировало работу эндогенных осцилляторов и приводило к генерации потенциалов действия разной амплитуды. Электрическая гетерогенность одноклеточных является отражением субстанциональной и функциональной изменчивости, и эти особенности определяют существование этой клетки как самостоятельного свободно плавающего организма.
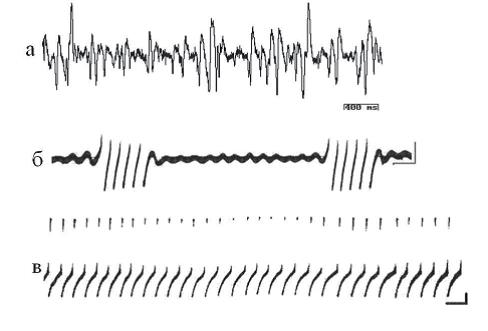
Рис. 4. Суммарная активность, зарегистрированная макроэлектродом в центральной области висцерального ганглия (а) и пейсмекерная активность изолированных нейронов моллюска Helix lucorum групповая (б) и регулярная (в). Калибровка: а – 400 мс, 50 мкВ; б, в – 1 c, 10 мВ.
Пейсмекеры улиток. Улитки (мы исследовали Helix pomatia и Helix lucorum) принадлежат к таксономической группе моллюсков, сформировавшейся в докембрии (более 650 млн лет назад). Все жизненно важные биологические функции многоклеточных организмов представляют собой ритмические процессы. Они осуществляются при помощи эндогенной осцилляторной активности нейронов, функции которой в нервной системе многоклеточных организмов весьма разнообразны: они отвечают за частоту сокращений сердца и дыхания, регулируют температурную чувствительность, контролируют деятельность желудочно-кишечного тракта, организуют электрическую ритмическую активность мозга, ответственны за проявление циркадианных ритмов и организацию двигательной активности, участвуют в создании энграмм памяти (Греченко, 2008). Спонтанные потенциалы действия, которые развиваются в клетках без очевидного синаптического возбуждения, были обнаружены у ряда нейронов позвоночных и беспозвоночных. Считается, что эти потенциалы действия возникают благодаря пейсмекерному механизму, который присущ данной клетке и является независимым от других нейронов. Существование эндогенной осцилляторной активности у нейронов моллюсков убедительно доказано работами Альвинг, Баумгартена и др., именно у улиток были впервые обнаружены ритмически генерирующие клетки (Arvanitaki, Chalazonitis, 1964). Опыты на полуинтактном препарате и на полностью изолированных клетках показали, что пейсмекерная электрическая активность характерна для многих нейронов ЦНС моллюсков, а функциональное разнообразие задач, в которые вовлекаются такие нейроны, исключительно велико (Литвинов, 1975). Клетки пейсмекерного типа участвуют не только в регуляции функциональных состояний, зависящих от времени суток, но и в осуществлении различных движений в качестве командных единиц. В висцеральном ганглии аплизии примерно 50–75% крупных нейронов демонстрируют спонтанные пейсмекерные разряды. Пейсмекерные разряды могут быть регулярными (ритмичный пейсмекер) или же состоять из периодических разрядов, разделенных периодами (рис. 4, б, в). С небольшими вариациями идентифицированные клетки проявляют тенденцию демонстрировать одну и ту же форму разрядов от препарата к препарату. Регистрация суммарной электрической активности (рис. 4, а) показывает существование частот в диапазоне от 0,5 Гц до 45 Гц. Поведение улиток сложное, они достаточно легко обучаются, что привлекает к ним внимание исследователей (Жизнь животных, 1968).
Обсуждение
1. Наши опыты показали, что осцилляции, выраженные на языке электрических процессов, являются самой древней формой электрической активности – они обнаружены у цианобактерий, биологический возраст которых почти равен геологическому возрасту Земли – не менее 3,5 млрд лет.
2. У цианобактерий разных видов – Oscillatoria terebriformis, Geitlerinema sp. и Halothece sp. от индивидуальных клеток зарегистрированы осцилляции постоянной частоты (2–3 Гц), которая оставалась неизменной в течение нескольких часов. Регистрация электрической активности макроэлектродом в сообществе цианобактерий показывает синхронизированные квазисинусоидальные ритмы частотой от 3–5 Гц до 30–40 Гц.
3. У микроорганизмов, ведущих одиночный, независимый от конспецификов образ жизни – как, например, у инфузории Paramecium caudatum – от каждого индивидуума зарегистрировано множество осцилляторов, каждый из которых характеризуется особой частотой, амплитудой, паттерном и формой колебаний. Свободно живущие подвижные микроорганизмы имеют разнообразную осцилляторную электрическую активность.
4. Такие сложные существа, как моллюски имеют большое количество нервных клеток, характеризующихся наличием пейсмекерной активности. У моллюсков Helix pomatia и Helix lucorum, имеющих нервную систему, разнообразие частотных характеристик электрических осцилляций пейсмекерных нейронов чрезвычайно велико – от 0,1 Гц до 10 Гц. Поведение Helix pomatia и Helix lucorum – сложное и разнообразное.
Синхронизация индивидуальных ритмов необходима для формирования социальной структуры. Способ выживания (частично назависимый или только в сообществе) определяется особенностями осцилляторной активности – это вывод, который можно сделать на основании результатов опытов на живых существах разного эволюционного уровня. Цианобактерии и дрожжи (грибы) ведут малоподвижный образ жизни. Чтобы выжить в агрессивной среде, цианобактерии формируют сообщества разной степени связности (биопленки, цианобактериальные маты), а дрожжевые клетки остаются относительно независимыми, но при этом формируют большую группу. Это позволяет таким организмам синхронизировать свои осцилляторы и осуществлять целенаправленное поведение. Когда деятельность членов сообщества направлена на решение жизненно важной задачи (например, освоение богатых пищей территорий, противостояние другому сообществу), то электрическая активность многих членов сообщества синхронизируется и это выражается в генерации высокоамплитудных ритмических колебаний суммарного потенциала. Осцилляции, зарегистрированные из зон, отличающихся активностью микроорганизмов, различны по частоте и амплитуде синхронизированной суммарной активности. Чем менее активен локус биопленки или мата, тем меньше выражена синхронизация осцилляций по частоте и тем ниже амплитуда колебаний. Эти результаты подтверждаются данными, полученными на других микробных пленках. Используя мультиэлектродную систему, имеющую высокое разрешение, исследовали электрическую активность двух видов микроорганизмов, строящих пленку, и одного – не создающего таковой. Частота потенциалов действия, зарегистрированных во время строительства пленки бактериального роста, показала одновершинный максимум с длительным последействием, что соответствует самому высокому уровню развития пленки. Такой пик не наблюдался в микробной среде, не производящей пленки (Masi et al., 2015). У многоклеточных организмов клетки – носители генетически заданной пейсмекерной программы – становятся структурообразующими элементами, формирующими вокруг себя систему синаптически связанных клеток, от их собственных «врожденных» свойств зависят многие особенности создаваемых нейронных систем, а следовательно, и поведенческих особенностей организма. Каждый осциллятор реализует определенную поведенческую программу. С повышением сложности внутриклеточных коммуникаций происходит и увеличение числа пейсмекеров, проявляющихся в электрической активности и поведении микроорганизма (примером могут быть парамеции). Сложность поведения зависит от числа осцилляторов, которые определяют свободный (относительно независимый) способ выживания.
Характеристики осцилляций влияют на развитие социальности. Прокариоты демонстрируют разнообразное поведение, предполагающее информативные коммуникационные связи и имеют сложные сигнальные механизмы для адаптации к изменениям температуры, осмотического давления, pH, количеству питательных веществ и другим существенным для их жизни свойствам окружающей среды. Одним из механизмов межклеточного взаимодействия прокариот является система, состоящая из белкового рецептора, чувствительного к определенному сигналу, и регулятора ответа, который контролирует экспрессию нужных генов (Смольская, Песнякевич, 2006). В данном случае контроль регуляции генов осуществляется посредством небольших диффузионных молекул-феромонов (Duan et al., 2009). Такие сигнальные молекулы позволяют индивидуальным бактериальным клеткам начать совместное действие тогда, когда будет достигнута пороговая плотность бактериальной популяции (кворум) (Lyon, 2007). Как показывают эксперименты, во время формирования колоний бактерии генерируют множество сигналов разной физической природы, среди которых весьма существенную роль играют электрические осцилляции, характеризующие определенные виды метаболических процессов этих существ (Masi et al., 2015).
В наших экспериментах показано, что цианобактерии имеют чрезвычайно устойчивую индивидуальную электрическую активность, для которой характерно постоянство периодичности. Цианобактерии выживают только в сообществе, и синхронизированная электрическая активность является пусковым механизмом совместной деятельности. Если по каким-то причинам синхронизации не происходит, члены сообщества погибают. Как показывают исследования, бактериальные биосоциальные системы характеризуются единым жизненным циклом, следовательно, их активность упорядочить во времени проще, чем, например, у дрожжей. У эволюционно более продвинутых дрожжевых клеток (одноклеточных эукариот – грибов) внутриклеточная регистрация выявляет наличие сложных паттернов эндогенной активности, среди которых трудно выделить электрические процессы, отражающие повторяющиеся ритмы. Это происходит также по причине множественности осцилляторов, имеющихся в каждой дрожжевой клетке.
Объединившись в колонии или пленки, живая система микросуществ ведет себя подобно многоклеточному организму или социальному сообществу повышенной сложности. В естественных условиях роста бактерии живут в иерархически построенных сообществах (Сумина, 2006). Принято считать, что основным регулятором жизнедеятельности сообщества является «чувство кворума» (ранее известное для бактериальных культур как «плотностная зависимость»), которым бактерии пользуются, чтобы инициировать изменение поведения в зависимости от плотности бактериального населения. Чтобы решить новую проблему, они оценивают ее через коллективное чувство, реализуют индивидуальную сохраненную информацию о прошлом опыте и затем осуществляют информационный процесс, распределенный по бактериям внутри колонии (109–1012 индивидуумов), создавая своеобразный «супермозг» (Ben-Jacob, 2003). Затем, благодаря механизму синхронизации, объединяют усилия для решения жизненно важной задачи. Этот процесс совершается путем морфологического усложнения колонии через иерархический временно-пространственный паттерн. Высокая сложность колонии обеспечивает степень пластичности и гибкости, требуемую для лучшей колониальной адаптации к динамичной окружающей среде. У прокариот, имеющих ограниченные биологические возможности, возникает своеобразное решение проблем, возникающих из-за их примитивного внутреннего устройства – они объединяются, создавая временный функционально специализированный «супермозг». Согласно нашим данным, работа такого образования основана на координированной синхронизированной осцилляторной активности членов сообщества.
Чем разнообразнее индивидуальные осцилляторы микроорганизма, тем меньше он нуждается в поддержке общества. Примером может быть свободноживущая инфузория, имеющая множество эндогенных осцилляторов. Дальнейшее развитие электрически выраженных осцилляций связано с появлением многоклеточных существ, нервная система которых содержит нейроны со встроенными генераторами (Соколов, 2003). Эти нейроны, выполняющие разнообразные функции, организуют и двигательную, и когнитивную деятельность высокоразвитых живых существ, включая человека (Данилова и др., 2013). Многоклеточные организмы координируют свою деятельность и развитие посредством сложных взаимодействий составляющих их клеток. Структурная и физиологическая сложность биопленки приводит к мысли о том, что она является координированной и кооперированной системой, аналогичной мультиклеточному организму. Микроорганизмы создают объединения, которые характеризуются электрической активностью, имеющей такие же частоты, как мозг человека (Греченко и др., 2013). Тогда можно предположить, что формирование колонии, пленки или мата происходит для того, чтобы создать «объединенный разум», достаточный для выработки стратегии поведения колонии в определенной жизненной ситуации. Отметим, что индивидуальные ритмы прокариот единообразны, мала вариативность частоты генерации.
У живых существ, имеющих сложную нервную систему, синхронизация осцилляторов существенна при выполнении действий, направленных на общие для участников цели. Например, в опытах на японских куропатках обнаружено, что если отклоняющихся по времени проявления активного поведения птенцов вводили в группу с отличающейся от них по времени активностью, то группа их отвергала (Formanek et al., 2011). Результаты экспериментов на людях показывают, что при взаимодействии участников во время решения задачи для достижения общей цели осцилляторная активность определенных областей мозга синхронизируется и ее вспышки ассоциируются с действиями партнеров (Funane et al., 2011). Эта активность по поводу достижения цели проходит на фоне синхронизированной активности определенных областей мозга каждого из участников. Следовательно, индивидуальная эндогенная ритмика предсказывает возможность социальной интеграции индивидуума