Введение
Широкое развитие программно-аппаратных решений, связанных с регистрацией тех или иных сигналов, генерируемых человеческим организмом, стало толчком к появлению большого класса приборов, обеспечивающих ввод команд пользователя в устройства, функционирующие на основе компьютеров, в самых различных вариантах: от классических настольных до интегрированных, например, в транспортные системы. Осуществляется конструирование новых эргатических (человеко-машинных) систем, и актуальной задачей в области создания аппаратно-программных средств является оценка эффективности их работы. Под эффективностью следует подразумевать точность и скорость выполнения команд, простоту освоения человеком системы, функциональное напряжение, возникающее как при обучении, так и при работе с эргатической системой. Одними из перспективных интерфейсов «человек—компьютер» с точки зрения увеличения пропускной способности как для обычного пользователя, так и для пользователя с ограниченными возможностями являются интерфейсы «мозг—компьютер», окулографические, дыхательные, миографические [Туровский, 2017; Lotte, 2007; Martin, 2011; McFarland, 2011; Zhang, 2016]. Очевидно, что оценку функционального напряжения пользователя необходимо осуществлять непосредственно в ходе работы с указанными интерфейсами; и наиболее оптимальным вариантом оценки состояния нервно-психического напряжения является вариабельность сердечного ритма (ВСР) [Бабунц, 2002; Баевский; Кулаичев, 2007] оценка которой должна проводиться с учетом индивидуальных психофизиологических особенностей человека, имеющих часто генетический характер.
Ранее нами было показано, что успешность освоения интерфейсов находится в тесной взаимосвязи с особенностями генотипа оператора, в частности, с полиморфизмами генов, которые участвуют в метаболизме серотонина [Turovsky, 2019]. Мы предполагаем, что параметры ВСР пользователей интерфейсов «мозг—компьютер», окулографических, дыхательных и миографических интерфейсов также могут зависеть от SNV (single nucleotide variation) в генах, которые так или иначе связаны с функционированием вегетативной нервной системы.
Целью данной работы являлось определение взаимосвязи между ВСР при работе с нейрокомпьютерными интерфейсами и SNV в генах, связанных с функционированием серотонинергической системы мозга, поскольку данная система наиболее обширно охватывает все отделы мозга, в том числе и гипоталамо-гипофизарную систему, которая во многом опосредует реакцию организма на стресс.
Интерес представляет ген TPH2 (tryptophan hydroxylase 2), который катализирует ключевую реакцию синтеза серотонина, и ген HTR2A (5-hydroxytryptamine receptor 2A), кодирующий 5-НТ2А-рецептор, способный одновременно оказывать возбуждающее и тормозящее действие в различных отделах головного мозга [Martin, 1998]. Дополнительно мы проанализировали мутации в генах APOE (apolipoprotein E), участвующих в метаболизме липидов и холестерина в крови и мозге, оказывающих значительное влияние на ВСР и связанных с возможным развитием артериосклероза и сердечно-сосудистых заболеваний [Puglielli, 2003; Ren, 2010].
Материалы и методы
Регистрация ВСР. В исследовании приняли участие 23 добровольца мужского пола в возрасте от 18 до 21 года. Работа осуществлялась в соответствии с этическими стандартами Хельсинкской декларации «Этические принципы проведения научных медицинских исследований с участием человека». Испытуемые проходили предварительный инструктаж и подписывали информированное согласие. Исследование одобрено этическим комитетом Воронежского государственного университета: протокол заседания № 42-03 от 13.02.2017. Все испытуемые ранее не имели опыта управления ни одним из интерфейсов, которые использовались в эксперименте. Регистрация ВСР осуществлялась прибором Поли-Спектр-12 (производство ООО «Нейрософт») при частоте дискретизации 1 кГц и включенном режекторном фильтре и фильтре дрейфа изолинии. Электроды фиксировались на конечностях испытуемого согласно стандартной схеме регистрации ЭКГ. В ходе эксперимента испытуемый располагался в кресле, заняв удобную для себя позу перед 21 LCD монитором, на котором демонстрировалась либо активная графическая форма соответствующего интерфейса (эксперимент, «работа») либо неактивная графическая форма соответствующего интерфейса (фоновая запись, «фон»). Последняя представляла собой пятиминутную регистрацию исходных значений ВСР, когда процедура управления и использование того или иного интерфейса не осуществлялись. Во всех экспериментах регистрация проводилась во второй половине дня, начиная с 14.00 часов, при этом после регистрации «фона» испытуемый переходил к работе с интерфейсом. Фоновая регистрация данных осуществлялась до эксперимента, во избежание остаточных эффектов, связанных как с необходимостью возврата вегетативной регуляции в исходное состояние, так и возможными модуляциями эмоционального фона, возникающими в ответ на выполнение или невыполнение задания. Несмотря на отсутствие возможности управления интерфейсом у испытуемых, все необходимые для управления датчики к пользователям были подключены, что обеспечило исключение возможности влияния их низкой эргономики на функциональное напряжение пользователя.. При обработке результатов ВСР, после удаления артефактов, анализировались параметры среднего значения RR-интервала (М); среднеквадратического отклонения (а); индекса вегетативного равновесия (ИВР) [Heart rate variability, 1996]; значения спектральной плотности мощности (СПМ), полученные на основе преобразования Фурье в стандартных частотных диапазонах [Кулаичев, 2007]; результаты вейвлет-преобразования исходного сигнала в стандартных диапазонах, позволяющих выделить тонические и нестационарные элементы («выбросы», отражающие активные регуляторные влияния) на кривой ВСР.
Задача испытуемых состояла в наборе десяти букв для каждого из интерфейсов, выбранных случайным образом. Перед началом эксперимента испытуемый не знал последовательность букв, которые ему надо будет набрать с использованием интерфейсов. Сразу после набора одной буквы испытуемому предъявлялась следующая.
Дыхательный интерфейс. В ходе работы с дыхательным интерфейсом (ДИ) испытуемый должен был в эксперименте А, изменяя скорость и объем выдоха, добиваться изменения положения по оси Y курсора, перемещающегося в течение 8 секунд от левого края экрана к правому. В правой области экрана располагалось 6 блоков букв. При совмещении курсора с соответствующим блоком осуществлялся первый уровень выбора требуемого символа. Буквы выбранного блока после этого выбора формировали новый ряд из 6 блоков, каждый из которых содержал теперь только одну букву. Перемещение курсора начиналось заново, что в итоге обеспечивало выбор конкретного символа из представленных (рис. 1а).
В эксперименте Б круговое поле графического интерфейса было разбито на 6 секторов, каждый из которых был активен в течение 2 секунд, о чем пользователь информировался изменением цвета соответствующего сектора (рис. 1б). Для выбора необходимого сектора пользователю требовалось совершить интенсивный выдох. Превышение порога звукового давления на мембрану микрофона фиксировалось как выбор сектора испытуемым. Символы, находившиеся в нем, автоматически перемещались на другие, предварительно очищенные сектора. Таким образом, для выбора символа, как и в предыдущем случае, необходимо было два раза подряд правильно выбрать тот или иной блок.
Аппаратной частью электромиографического интерфейса (ЭИ) служил электроэнцефалограф «Нейрон-Спектр-4 ВП» производства ООО «Нейрософт». Электроды располагались в проекции плечелучевой мышцы на 7—10 см дистальнее латерального мыщелка плечевой кости. Межэлектродное расстояние составляло 4—5 см. Полиграфические каналы прибора обеспечивали регистрацию ЭМГ с частотой до 40кГц, которая затем потом программно снижалась до 500 Гц. Дизайн графического интерфейса полностью соответствовал дизайну графического интерфейса, использующегося в эксперименте ДИ серии Б. Испытуемый должен был напрягать мышцы предплечья в случае, если подсвеченный сектор на графическом интерфейсе соответствовал блоку букв или букве, которую необходимо было выбрать.

Работа окулографического интерфейса (ОИ) основывалась на анализе видеоизображения зрачка, положение которого преобразовывалось в положение курсора на форме программы (таким образом осуществлялась обратная связь «компьютер—человек»). Задача испытуемого состояла в помещении курсора под контролем зрения на один из блоков, содержащий требуемую для выбора букву (рис. 2). После этого остальные пять блоков очищались от букв, а шесть букв выбранного блока разделялись между шестью имеющимися блоками так, что в каждом осталось по одной букве. Дальнейший выбор осуществлялся аналогично.

Работа нейрокомпьютерного («мозг—компьютер») интерфейса (НКИ) основывалась на детекции устойчивых зрительных вызванных потенциалов (SSVEP). Регистрация ЭЭГ осуществлялась при помощи комплекса «Нейрон-Спектр-4 ВП» производства ООО «Нейрософт» с включенным режекторным фильтром и выключенными фильтрами высоких и низких частот. Фотостимуляция осуществлялась шестью диодами белого цвета (0,5 Вт), расположенными на специальной рамке по краям монитора. Частота стимуляции составляла 9.009, 10.10, 11.11, 12.19, 13.33, 14.49 Гц. Регистрация ЭЭГ активности осуществлялась электродами в позициях O1, O2, Oz, P3, P4, Pz; референтным электродом служил объединенный ушной электрод. Логика изменения графического интерфейса соответствовала таковой для окулографических исследований.
Точность работы всех интерфейсов определялась по числу правильно выбранных блоков, содержащих необходимый символ, заданный испытуемому.
Генотипирование. Выделение ДНК из буккального эпителия проводилось с использованием набора «Проба-ГС» (ДНК-Технология, Россия). Генотипирование проводилось методом ПДРФ-анализа. ПЦР проводили на приборе Bio-Rad CFX96TM (Bio-Rad, USA) с использованием набора qPCRmix-HS (Evrogen, Russia). Общую денатурацию проводили при температуре 95°С в течение 3 мин; денатурация в начале цикла — 95°С, — 30 с; отжиг праймеров — 59—71°С, 30 с, элонгация — 72°С, 30 с; количество циклов — 35. Информация об используемых праймерах и эндонуклеаз рестрикции представлена в табл. 1.
Таблица 1
Характеристики используемых праймеров и эндонуклеаз
|
№ |
SNV |
Праймеры |
Эндонуклаза рестрикции |
Размер продукта |
|
1 |
rs6313 |
F: TGAGCTCAACTACGAACTCCCTA R: AGAGACACGACGGTGAGAGG |
Msp I |
T/T — 172 C/C — 99; 72 C/T — 172; 99; 72 |
|
2 |
rs429358 |
F: CGCCTCGCCTCCCACCTGAGCAAG R: CGCTCGTCGCCCTCGCGGG |
HspA I |
T/T — 72 C/C — 45; 27 T/C — 72; 45; 27 |
|
3 |
rs4570625 |
F: GGCTAAATTGAACCCTTACCTTT R: GGTAATCAAGATATCCATTGCC |
Psi I |
G/G — 301 T/T — 89; 212 G/T — 89; 212; 301 |
|
4 |
rs4290270 |
F: TTTTGTTTTGGGTGCCATTT; R: TGCATGGGAAGGGTATTTTC |
FauND I |
T/T — 209 A/A — 134; 76 T/A — 209; 134; 76 |
В статистическом анализе полученных результатов использовались методы дескриптивной статистики. Учитывая, что распределения большинства показателей не являются нормальными, применялись критерии непараметрической статистики: критерии Краскела—Уоллиса, Манна—Уитни, Фридмана и Вилкоксона для парных случаев [Гланц, 1998; Рунион, 1982], при этом параметр а принимался равным 5%. Учитывался эффект множественных сравнений. Обработка осуществлялась как в программном пакете Stastistica 8.0 так и в оригинальном ПО. Конкретная версия пакета Stastistica 8.0 верифицировалась тестовыми выборками с известными результатами обработки.
Результаты и обсуждение
Показатели ВСР при работе с интерфейсами «человек—компьютер».
Результаты проведенного анализа SNV rs4570625 гена TPH2, который катализирует лимитирующую реакцию синтеза серотонина, указывают на единственное различие: при работе с миографическим интерфейсом показатели вегетативного равновесия (индекс, рассчитанный по Р.М. Баевскому) для генотипа G/G были значимо (р<0,033, критерий Манна—Уитни) ниже, чем для генотипа G/Т. Однако это наблюдение является единичным, как для исследуемой мутации, так и для исследуемого интерфейса, что не позволяет сделать выводы о её возможном влиянии на исследуемый процесс.
Взаимосвязь между показателями ВСР и SNV rs6313.
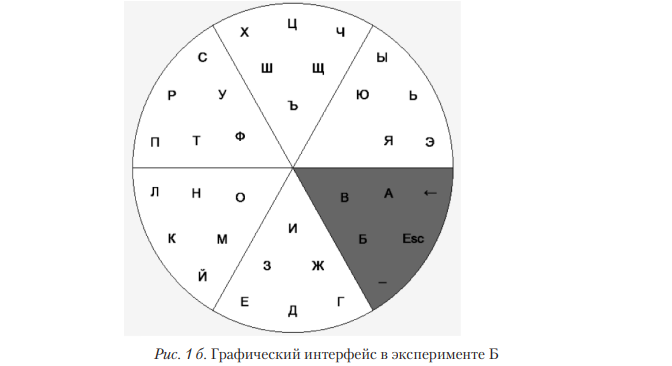
Взаимосвязи между SNV rs6313 и показателями ВСР при работе с различными интерфейсами выявили широкий спектр различий. Как известно, показатель SD отражает суммарную активность управляющих влияний на вариабельность сердечного ритма. Таким образом, гомозиготы С/С имели больший размах регуляторных воздействий (рис. 3) для окулографического интерфейса. При этом был выявлен более высокий уровень моды квадратов коэффициентов вейвлет-преобразования значений этого же генотипа в LF-частотном диапазоне (рис. 4), связываемого либо с симпатическими влияниями, либо с активностью сосудодвигательного центра [Рябыкина; Schächinger, 2001]. Модальные значения в аспекте анализа ВСР можно интерпретировать как тоническую активность управляющих центров (симпатических или сосудодвигательных). Генотип С/С в SNV rs6313 ассоциирован с увеличением экспрессии 5’-нетранслируемой области гена HTR2A, что приводит к увеличению эффективности трансляции этого гена и увеличению концентрации белка 5-HT2A-рецептора [Smith, 2013]. Изменение экспрессии HTR2A в ядрах одиночного пути (лат. nucleus tractus solitarii) участвует в вегетативной регуляции сердечного ритма и кровяного давления [omet, 2007]. Также значимые различия были выявлены при работе с НКИ: значения SD у лиц с генотипом С/T значимо уступали пользователям, у которых был выявлен генотип С/С (50±5мс; 84±16мс; p=0,035, критерий Краскела—Уоллиса). Таким образом, как и в случае с окулографическим интерфейсом, выявлена связь СС-гомозиготы с более выраженными влияниями на вегетативное управление ВСР в результате увеличения экспрессии гена HTR2A [Smith, 2013]. Аналогично в несколько раз различались и показатели частотно-временной картины ВСР, полученной на основе вейвлет-преобразования. Для С-аллеля характерны более высокие показатели тонических влияний на ВСР, представленных модальными значениями квадратов коэффициентов вейвлет-преобразования (80223±20798 мс2 для Т-аллеля и 250265±83429 мс2 для С-аллеля). Суммарная мощность так называемых «выбросов» — областей, где структура ВСР в исследуемом частотном диапазоне резко изменялась, отражая активные регуляторные воздействия, — также отличалась более высокими значениями в случае С-аллеля, чем в случае Т-аллеля: 349669±152920 (нижний квантиль 98042, мода 243551, верхний квантиль 377323) мс2 против 175573±75941 (нижний квантиль 53745, мода 87049, верхний квантиль 120335) мс2 соответственно (р=0,033, критерий Краскела—Уоллиса).
Взаимосвязь между показателями ВСР и SNV rs4290270.
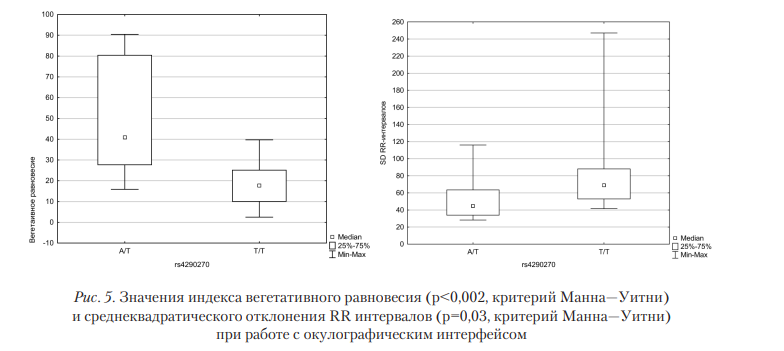
Наибольшее количество различий в показателях вегетативного равновесия было обнаружено при анализе взаимосвязи между ВСР и SNV rs4290270 гена TPH2 (табл. 2). Выявленные изменения подтверждаются и классическими методами анализа ВСР. Так, индекс вегетативного равновесия для Т/Т-генотипа значимо (р<0,05) ниже, чем для гетерозиготного носителя (рис. 5).
C учетом того, что не было обнаружено различий в «фоновых» значений в серии экспериментов с окулографическим интерфейсом в зависимости от точечных мутаций, следует признать, что работа с окулографическим интерфейсом приводит к существенному усилению регуляторного влияния парасимпатического отдела ВНС для генотипа Т/Т.
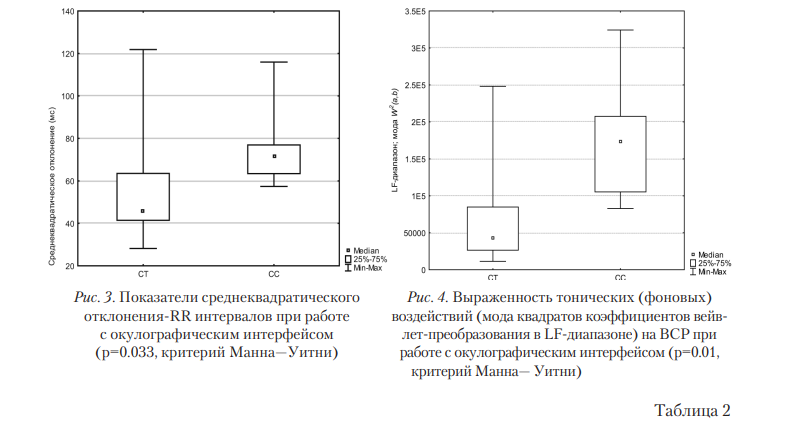
Таблица 2
Значения результатов вейвлет-преобразования HF ВСР при работе с окулографическим интерфейсом (M±m; Q25, Me, Q75, мс2)
|
Генотип |
мода (W2(a,b)) (р=0,026) |
Среднеквадратическое отклонение (W2(a,b)) (р=0,035) |
«Выбросы» (W2(a,b)) (р=0,035) |
|
A/T |
1685,99±690,53; 368; 1004; 1575 |
1218,85±536,14; 352; 686; 1385 |
2430,66±1275,28; 727; 1138; 1778 |
|
T/T |
10127,84±6527,050 1101; 3334; 5804 |
26029,40±22586,53 862; 3000; 4453 |
63098,64±55741,16; 1790;4836; 10131 |
Примечание: сравнение проведено по критерию Манна—Уитни.
При этом повышаются как тонические влияния (мода (W2(а,b))), так и активное регулирующие воздействие («Выбросы» (W2(а,b))).
Так, результаты анализа свидетельствуют о более низких значениях индекса вегетативного равновесия в случае Т/Т-генотипа и в то же время о более высоких значениях среднеквадратического отклонения RR-интервалов (рис. 5). При анализе результатов работы с ДИ были также выявлены разные паттерны ВСР в зависимости от выявленных точечных мутаций (табл. 3). Как видно из табл. 3, при изменении режима дыхания, необходимого для работы с ДИ, в случае Т/Т определяются существенно более высокие значения активности как симпатического, так и парасимпатического отделов ВНС. При этом прослеживается та же динамика вегетативной активности, что и при работе с ОИ: наблюдается рост как тонических влияний, так и активных регуляторных воздействий. Однако в отличие от ОИ изменения затрагивают и симпатический отдел ВНС (или сосудодвигательный центр), что, вероятно, объясняется существенно большим влиянием изменения режима дыхания при работе с ДИ в отличие от вариантов ОИ, НКИ и ЭИ. Такие результаты могут объясняться изменением уровня серотонина, который, как известно, играет ключевую роль в регуляции вегетативной нервной системы [Alenina, 2009].
При этом уровень серотонина напрямую зависит от экспрессии гена TPH2, белок которого катализирует лимитирующую реакцию синтеза серотонина [Chen, 2013]. T-аллель в SNV rs4290270 ассоциирована с более высоким уровнем экспрессии гена TPH2. Это может приводить к увеличению уровню серотонина в головном мозге и активации как симпатического, так и парасимпатического отделов ВНС.
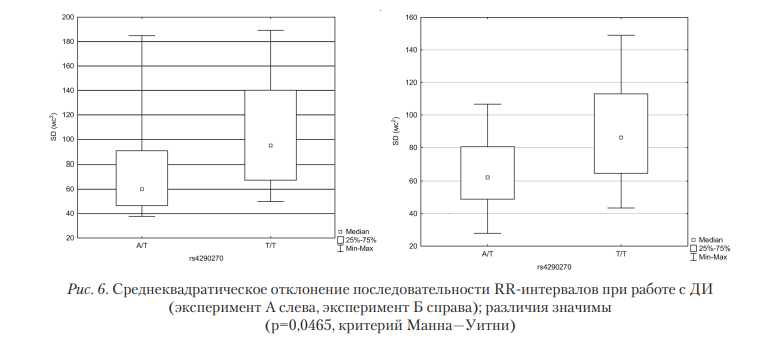
Итак, результаты анализа ВСР указывают на одинаковую динамику показателей среднеквадратического отклонения для различных генотипов, как в эксперименте А, где испытуемым было необходимо поддерживать уровень выдыхаемого воздуха в определенном диапазоне, так и в эксперименте Б. При анализе генотипа Т/Т обнаруживается больший разброс кардиоинтервалов (рис. 6), что можно интерпретировать как большую выраженность регуляторных влияний ВНС, находящейся в тесной взаимосвязи с изменением уровня серотонина [Jordan, 2005]. Схожие различия продемонстрированы и в результатах, полученных на основе спектрального анализа. При отсутствии различий в HF-диапазоне при разделении групп по генотипу обнаруживаются значимо большие значения СПМ LF-диапазона в случае T/Т-генотипа. Таким образом, при работе с дыхательным интерфейсом вне зависимости от характера этой работы (поддержание воздушного потока на нужном уровне или короткие и интенсивные его изменения) в первую очередь изменяются параметры частотного диапазона, связанного с симпатическим отделом ВНС, влияния которого и обеспечивают серотонин-индуцируемые изменения ВНС. При анализе динамики результатов вейвлет-преобразования между фоновым состоянием пользователя и работой с ДИ получены результаты, представленные в табл. 4. Во всех экспериментальных пробах наблюдается увеличение как тонических влияний на ВСР, так и активных регуляторных воздействий. Анализ спеицифики работы с НКИ (ИМК) позволил выявить изменения, связанные с активностью симпатического отдела ВНС. В случае Т/Т-генотипа показатели активности симпатического отдела ВНС являются более высокими (p<0,046); при этом в ходе работ с интерфейсом носители этого генотипа в значительно меньшей степени (р<0,03) реагировали снижением СПМ в LF диапазоне, чем носители гетерозиготы.
Значения результатов вейвлет-преобразования ВСР при работе
с дыхательным интерфейсом (M±m; Q25, Me, Q75; мс2)
|
rs 4290270 |
Эксперимент А |
Эксперимент Б |
|||
|
А/Т |
Т/Т |
А/Т |
Т/Т |
||
|
HF-диапазон |
мода (W2(a,b)) |
2354±652; 950; 1452; 3734 (р=0,003) |
16168±6830; 2375; 8694; 13421 |
2574±948; 580; 1051; 4244(р=0,016) |
9576±2565; 3042; 5935; 15234 |
|
a (W2(a,b)) |
2821±919; 687; 1613; 5962(р=0,03) |
28520±19372 2049; 7396; 15841 |
3301±1559; 550; 871; 4406 (р=0,016) |
11988±3466; 2639; 5608; 21755 |
|
|
«Выбросы» (W2(a,b)) |
5946±2151; 1226; 3095; 10180 (р=0,04) |
66639±47761; 3851; 15422; 30833 |
7157±3807 1165; 1496; 8667 (р=0,01) |
25550±7924; 5318; 11355; 31530 |
|
|
LF-диапазон |
мода (W2(a,b)) |
133902±25201; 92481; 106347; 207698 (р=0,03) |
284038±53946135308; 208708; 358422 |
84010±16012** 38834; 77606; 135284 (р=0,0006) |
224072,7± 28722,36; 160350; 198282; 263004 |
|
a (W2(a,b)) |
100088±17082; 60029; 96225; 163338 (р=0,008) |
320693±97157121506; 168163; 410315 |
72446± 1647324772; 63143; 125536(р=0,0004) |
225033,7± 35693,52; 157895; 181485; 204559 |
|
|
«Выбросы» (W2(a,b)) |
173234± 35103; 87216; 145774; 264184 (р=0,005) |
721502± 228108; 244895; 324878; 927691 |
137834±36412 48731; 109861; 208617 (р=0,001) |
441853±80579; 279109; 320384; 418179 |
|
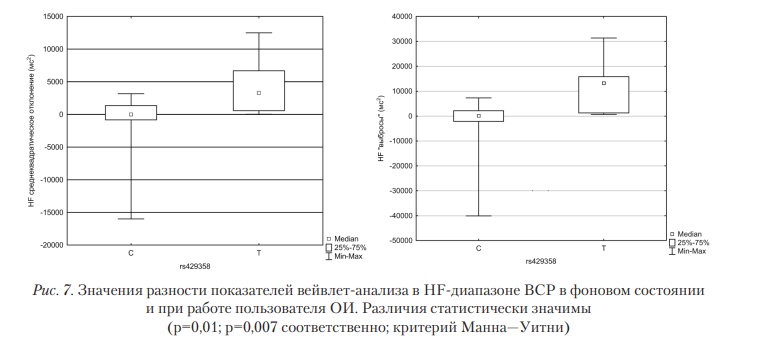
Взаимосвязь между показателями ВСР и SNV rs429358.
В данном случае различия выявлены для SNV rs429358 (рис. 7). В случае С-мутации работа с ОИ приводит к росту активности регуляторных парасимпатических влияний, при этом тонические парасимпатические влияния сохраняются неизменными. Ранее было показано, что мутации в генах, участвующих в метаболизме липидов, в частности в гене APOE, могут оказывать влияние на параметры ВСР, однако непосредственной связи с SNV rs429358 выявлено не было [Ren, 2010]. В целом можно отметить, что при работе с ОИ именно парасимпатические влияния претерпевают наиболее значимые изменения, различающиеся в зависимости от генотипа пользователя
Таблица 4
Разность показателей вейвлет-анализа ВСР в фоновом состоянии пользователя и при его работе с ДИ (эксперимент А) в зависимости от мутации rs 4290270.
(M±m; Q25, Me, Q75; мс2)
|
Генотип |
HF диапазон |
||
|
мода (W2(a,b)) |
с (W2(a,b)) |
«Выбросы» (W2(a,b)) |
|
|
A/T |
-729±717 -2084; -472; -25 |
-708±991 -1265; -273; 21 |
-1219±2303 -1845; -404; -23 |
|
T/T |
-12891±6888-10413; -4450; -1990 (р=0,007) |
-24281±19549 -9362; -2310; -276 (р=0,03) |
-57297± 48223 -18504; -4853; -1028 (р=0,046) |
Примечание: сравнение по критерию Манна—Уитни.
.
Заключение
В работе проведен анализ взаимосвязи SNV в генах HTR2A, APOE и TPH2 с показателями ВСР при освоении пользователями ряда интерфейсов «человек—компьютер»: «мозг—компьютер», окулографических, миографических и дыхательных. Результаты анализа свидетельствуют о том, что в ряде случаев определяется взаимосвязь SNV с особенностями вегетативной регуляции вариабельности сердечного ритма, заключающаяся как в изменении тонических воздействий, так и характеристик кратковременных воздействий на ВСР. С-аллели полиморфизма rs6313 гена HTR2A характеризуются более высокими показателями тонических влияний на ВСР, что, вероятно, связано с увеличением уровня белка серотонинового рецептора, который участвует в вегетативной регуляции сердечного ритма. Генотип Т/Т SNV rs4290270 гена TPH2 связан с большим разбросом кардиоинтервалов.
Такого рода взаимосвязь может объясняться увеличением экспрессии гена TPH2, которое, в свою очередь, приводит к увеличению уровня серотонина в головном мозге вследствие активации как симпатического, так и парасимпатического отделов ВНС. Результаты проведенного исследования позволяют расширить представления о генетической детерминированности уровня функционального напряжения при освоении перспективных интерфейсов «человек—компьютер».