Пластичность традиционно рассматривается как основа мнемических, когнитивных и семантических и других психических процессов. В психофизиологии исследование мембранных механизмов пластичности начали проводиться в 70-х годах ХХ в. Это стало возможным благодаря развитию экспериментальной и компьютерной техники. На кафедре психофизиологии под руководством профессора Е.Н. Соколова исследования пластичности проводились на нейронах высших и беспозвоночных животных. В результате полученных данных и анализа литературы Е.Н. Соколов предложил модель концептуальной рефлекторной дуги (КРД) для описания взаимодействия между нейронами при осуществлении поведенческого акта. В состав КРД были включены несколько типов нейронов, выполняющих определенные функции в анализе внешних сигналов и реализации поведенческого акта: рецепторы, предетекторы, детекторы, модулирующие, командные и двигательные. Рецепторы преобразуют энергию внешних воздействий в нейрональный код, необходимый для дальнейшего анализа сигнала. Предетекторы и детекторы селективно выделяют параметры действующего сигнала и передают информацию к командным нейронам. Командные нейроны определяют осуществление целостной поведенческой реакции или ее отдельного фрагмента и управляют набором мотонейронов, возбуждающих двигательные единицы. Модулирующие нейроны самостоятельно не вызывают реакций, они перераспределяют приоритеты реакций командных нейронов на основании внешних и внутренних влияний. В концептуальной рефлекторной дуге выделяются три основные части: сенсорная (рецепторы, предетекторы, детекторы), принятия решения (командные и модулирующие нейроны) и исполнительная (мотонейроны и мышцы) (Соколов, 1981).
В дальнейшем Е.Н. Соколов расширял и дополнял эту основную схему, включая новые типы нейронов и их связей для описания механизмов функционирования мозга при осуществлении разной сложности психологических процессов. Последовательное развитие этой концепции привело Е.Н. Соколова к созданию нового векторного представления о работе мозга человека и нейронных механизмах сенсорных, когнитивных и исполнительных процессов (Соколов, 1995). Этот подход объединяет все уровни психофизиологического анализа психических явлений от поведения животных до сознания человека, включая потребности, эмоции и функциональные состояния.
Нейронные механизмы пластичности традиционно рассматривались как два основных процесса: синаптический и эндонейрональный. Новые данные о механизмах пластичности были получены в работах Е. Н. Соколова и его сотрудников к концу 70-х годов. Наиболее значимым было изучение ритмоводящего пейсмекерного потенциала, наличие которого в нейронах превращает их из простых сумматоров синаптических воздействий в управляемые генераторы (Соколов и др., 1975).
Следующим шагом в исследовании пластичности стало использование идентифицированных командных нейронов оборонительного поведения виноградной улитки, имеющих латентный пейсмекерный механизм. В составе концептуальной рефлекторной дуги эти элементы выполняют функцию «принятия решения». Изучение механизмов пластичности на препаратах нервной системы беспозвоночных животных соответствовало представлениям об идентичном строении мембран нервных клеток независимо от эволюционного уровня развития организма. В конце 70-х – начале 80-х годов исследование пластичности командных нейронов перешло на следующий уровень – мембранный. Для этого использовался оригинальный метод последовательной замены физиологического раствора, окружающего препарат изолированной ЦНС и полностью изолированных КН.
Путем последовательной замены и инкубации препарата в физиологических растворах (бескальциевый – замена хлорида кальция на хлорид кобальта, кадмия или марганца; безнатриевый – замена хлорида натрия буфером и безнатриевый+ бескальциевый – замена хлорида натрия буфером и хлорида кальция хлоридом кобальта, кадмия или марганца) изучались реакции потенциалзависимых ионных каналов мембраны нейронов и пейсмекерных потенциалов (Хлудова, 1989). Путем последовательной замены и инкубации препарата в физиологических растворах разного состава проводилось тестирование электрической чувствительности командных нейронов при действии внутриклеточных деполяризационных импульсов разной интенсивности в восходящем и нисходящем порядке (рисунок 1).
Это исследование показало, что генез пейсмекерного потенциала нейронов обеспечивается натриевой и кальциевой проводимостью мембраны (норма), а также при исключении одной из них в эксперименте. Инактивация кальциевой проводимости мембраны командного нейрона увеличивала латентный период появления первого спайка и снижала амплитуду потенциалов действия. У многих нейронов усиление деполяризующих импульсов внутриклеточного тока приводило к увеличению амплитуды и частоты пейсмекерных колебаний. Пейсмекерные волны проявляются перед возникновением потенциалов действия или в середине дейст
вующего стимула. При этом разряд командного нейрона смещается к концу действующего стимула. В растворе, блокирующем кальциевую проводимость мембраны, происходит сужение диапазона интенсивностей деполяризационных импульсов, вызывающих потенциалы действия в командных нейронах. Перевод препарата в безнатриевый раствор и его инкубация в течение 5–10 минут после отмывки физиологическим раствором нормального состава не изменяет характер разряда командного нейрона. Порог появления потенциалов действия, их амплитуда и частота, а также латентный период совпадают с этими параметрами в нормальном физиологическом растворе. Диапазон эффективных интенсивностей деполяризующего тока, вызывающих реакцию командного нейрона, совпадает с диапазоном для физиологического раствора нормального состава. Инкубация командного нейрона в безнатриевом растворе, содержащем хлорид кобальта, кадмия или марганца, приводит к полному отключению генерации потенциалов действия и пейсмекерных колебаний. Использование высоких интенсивностей деполяризующего тока не могло вызвать импульсации командного нейрона. Общее сопротивление мембраны нейрона при изменении состава растворов, омывающих препарат, не изменялось.
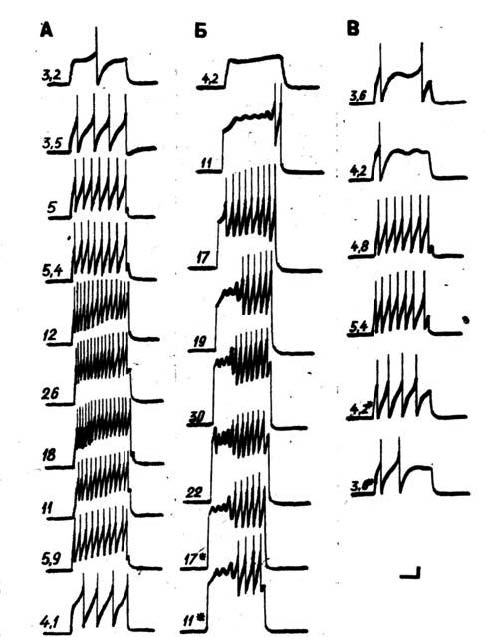
Рис. 1. Нейрограмма нейрона ЛПа 2 № 28 А – нормальном физиологическом растворе; Б – в растворе, содержащем 15 мМ хлорида кобальта; В – в безнатриевом растворе. Цифрами отмечена интенсивность деполяризационных стимулов; звездочками показаны интенсивности, совпадающие при повышении и снижении деполяризационных стимулов. Калибровка: 10 мВ; 1 с
Последовательная инкубация интактных и изолированных командных нейронов в физиологических растворах различного ионного состава показала, что генерация потенциалов действия в них полностью определяется натриевой и кальциевой проводимостью мембраны. Пейсмекерные потенциалы могут быть как натриевой, так и кальциевой природы. При последовательной функциональной изоляции каждого вида проводимости пейсмекерные потенциалы сохраняются, изменяется только порог их появления при действии внутриклеточного деполяризующего тока. Инактивация кальциевой проводимости снижает возбудимость командного нейрона. Исключение натрия как носителя проводимости не вызывает изменений возбудимости по сравнению с нормальным раствором.
Электрическая активность командных нейронов обеспечивается не только натриевой, но и кальциевой проводимостью мембраны, которая представлена ионными каналами с низкими и высокими порогами возбуждения. В целом электрогенез командных нейронов имеет смешанный характер, и определенный вклад в него вносит пейсмекерный потенциал. Функционирование пейсмекерного механизма в командных нейронах может быть обеспечено как медленным входящим кальциевым током, так и натриевым током в сочетании с медленным выходящим калиевым током. Проверка этих результатов была проведена при исследованиях свойств мембраны полностью изолированных командных нейронов совместно с Т. Н. Греченко. Вклад ионных проводимостей в пейсмекерный механизм и электрогенез командных нейронов полностью подтвердился и при изоляции нейронов из нервной системы (Хлудова, Греченко, Соколов, 1986). Пластические изменения в реакциях командных нейронов в составе ЦНС и полностью изолированных тестировались многократными применениями внутриклеточного деполяризационного импульса постоянной амплитуды (рисунок 2).
В этих условиях пластичность командного нейрона проявляется в виде снижения числа потенциалов действия при блокаде отдельно натриевого и кальциевого компонентов входящего тока и может определяться инактивацией пейсмекерного механизма. Пластичность пейсмекерных потенциалов представлена изменениями амплитуды и частоты по мере развития привыкания. Для отнесения постепенного снижения числа потенциалов действия к эффекту привыкания использовались различные экстрастимулы, приводившие к растормаживанию, т. е. восстановлению исходного ответа на деполяризующий ток. Эффект сенситизации пейсмекерного механизма командного нейрона может проявляться и как следствие замены физиологических растворов, что позволяет использовать этот прием для активации и тестирования пейсмекерных колебаний.
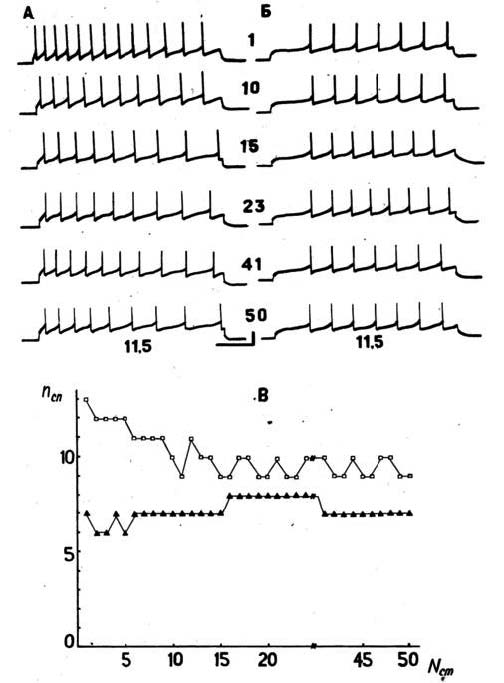
Рис. 2. Нейрограммы нейрона ППа 3 № 52:
А – в нормальном растворе и Б – в растворе, содержащем 15 мМ хлорида кобальта при интенсивности 11,5 нА; цифры отмечают номер стимула. Калибровка: 10 мВ, 1 с; В – графики привыкания нейрона ППаЗ в нормальном растворе (квадраты), в растворе, содержащем хлорид кобальта (треугольники). По оси абсцисс – номер стимула, по оси ординат – количество ПД
Таким образом, командные нейроны не утрачивают пластическую способность к привыканию и сенситизации, и это свойство прямо связано с вкладом пластичности ионных проводимостей мембраны в поведенческие реакции.