В начале был только ритм… Г. Бюлов
Периодичность процессов, обеспечивающая целостность и функционирование сложных систем, выявляется на всех уровнях существования живой и неживой природы. Так, геологическая история Земли характеризуется цикличностью протекания физических процессов: длина наиболее крупных циклов составляет 150–240 млн. лет, длина более мелких циклов составляет, по некоторым оценкам, 26–31 млн. лет (Raup, Sepkoski, 1986). В позднем протерозое, палеозое и кайнозое были выявлены эпохи оледенений длительностью от миллионов до десятков тысяч лет, чередовавшиеся с интерстадиальными, относительно более теплыми эпохами. Колебания сейсмической активности на Земле происходят с интервалом в 22–23 года. С притяжением Луны и Солнца связаны ритмические приливноотливные явления в гидросфере.
Особенно наглядно ритмика, представленная спектром гармоник с периодами от долей секунды до тысячелетий, проявляется на биогенных компонентах (Азроянц, 2003). Жизнь во всех ее проявлениях имеет циклический характер (Кондратьев, 1989; Чижевский, 1976). Среди факторов, способствовавших выживанию, важнейшую функцию выполняют ритмические события, которые разворачиваются в среде обитания и ассоциируются с внутренними процессами. Ритм – это повторение одного и того же события и воспроизведение одного и того же состояния через равные промежутки времени. В биологии под ритмичностью понимают периодические изменения интенсивности физиологических функций с различными периодами колебаний – от миллисекунд до нескольких лет. Так, все живые организмы имеют период покоя и активности, что согласуется с суточным движением и годовым обращением Земли. Сезонная ритмика проявляется в покое и вегетации растений, в линьке, спячке и миграции животных. В течение миллионов лет эволюции сформировалась суточная, сезонная и годичная ритмичность физиологических процессов. Все живые организмы, от одноклеточных до высших позвоночных, ориентируются во времени, поскольку обладают уникальным механизмом его измерения – биологическими часами, которые задают и контролируют ритм их жизнедеятельности. Функционирование такой системы обеспечивается путем периодического протекания таких важных процессов жизнедеятельности организма, как деление клеток, обмен веществ, питание, изменение уровня активности и др. Живые существа пользуются биологическими часами для того, чтобы регулировать поведение в соответствии с внутренним ощущением времени суток (Ralph et al., 2013). Функция биологических часов формируется в соответствии с суточным ритмом, который определяется в свою очередь периодическим вращением Земли вокруг своей оси, привносящим определенный ритм в смену времен года, ночи и дня, степени освещенности и температуры, атмосферного давления, влажности воздуха, космической и солнечной радиации.
Биологические часы регулируют большую часть физиологических и поведенческих проявлений жизни, включая сон, локомоторную активность, секрецию гормонов, метаболизм, формирование долговременной памяти и многие другие процессы (Загускин, 2010; Fernandez et al., 2003; Bell-Pedersen et al., 2005; Gillette, Sejnowski, 2005). Например, в опытах на позвоночных и беспозвоночных животных была показана циркадианная модуляция памяти: в экспериментах на морском моллюске аплизия с применением простых неассоциативных типов обучения (долговременную сенситизацию) (Fernandez et al., 2003) и более сложного ассоциативного оперантного обучения (Lyons et al., 2005) исследователи показали, что эффективность обучения регулируется циркадными ритмами. Сохранение функциональной зависимости между циркадными ритмами и обучением при формировании памяти у разных видов живых существ представляет собой важную основу для будущего анализа молекулярных механизмов, лежащих в основе сложного поведения.
Цикличность стимуляции, исходящей из внешней среды, и ее отражение в виде ритмики протекания внутренних процессов жизнедеятельности целостного организма способствовали формированию жизненного режима, основанного и регулируемого происходящими в определенное время и координированными с ритмикой внешней среды молекулярными взаимодействиями. В свою очередь постепенная гармонизация, координация внешних и внутренних событий и явлений способствовали ассоциированию молекулярных и клеточных процессов, результатом которого стала выработка соответствующих механизмов жизнедеятельности и форм поведения.
Можно предположить, что на ранних этапах жизни на Земле молекулярные изменения явились средством кодирования основной информации, необходимой для выживания: например, простые стимулы окружающей среды, такие как свет, температура, основные химические компоненты, требуемые для питания и утилизации биологической энергии, явились сигналами, запускающими метаболические химические реакции, которые, в свою очередь, стали основой осцилляторных явлений именно по причине циклического характера вращения Земли, определенной длительности прохождения ею пути вокруг солнца, а также цикличности поступления сигналов, исходящих из внешней среды и вызываемых этими природными факторами (Zhou et al., 2008). Повторяемость внешних событий сформировала циркадианные и цирканнуальные (сезонные) признаки, на основании которых протобионты формировали обеспечивающие регуляцию жизнедеятельности молекулярные механизмы. Такие ритмично повторяющиеся события могут рассматриваться в качестве одной из основ развития первобытной памяти – древнее живое вещество «запоминало», как нужно отвечать на циклически повторяющийся сигнал, какие биохимические реакции запускаются этим сигналом, и форму «правильного» поведения. Другими словами, биологические ритмы – это воспроизведение следов древней памяти на совпадающие во времени события внешней среды и структурно-молекулярные трансформации, запускаемые этими внешними сигналами. Ритмические явления характерны для жизнедеятельности живого организма любой степени сложности и любого эволюционного уровня – одноклеточных и многоклеточных прокариот, одноклеточных и многоклеточных эукариот – вплоть до высших растений и животных, включая человека.
В мировой научной литературе представлено множество данных, свидетельствующих о проявлении ритмичности в электрической активности различных элементов нервной системы живых существ разного эволюционного уровня, но практически отсутствуют экспериментальные данные об осцилляторной электрической активности у микроорганизмов. Основная задача нашей работы состояла в изучении электрической активности этих древнейших существ с целью выявления ритмических процессов, в которых реализуется воспроизведение энграммы первобытной памяти, у организмов разной степени сложности.
Метод
Опыты выполнены на наземных моллюсках Helix lucorum, инфузориях Paramecia caudatum, цианобактериях Oscillatoria terebriformis, дрожжах Saccharomyces cerevisiae и плесневых грибах Penicillium. Применялся метод регистрации электрической активности при помощи стеклянных микроэлектродов. Для работы с цианобактериями Oscillatoria terebriformis при меняли физраствор (в граммах на литр): NaHCO– 3, Na CO 17, K HPO– 0,5, NaCl – 30, KNO – 2,5, MgSO – 0,2, CaCl – 0,04, FeSO – 0,01. Электроды заполняли 1 М раствором KCl.
В опытах на дрожжах Saccharomyces cerevisiae использовалась культура «диких» дрожжей, помещенных в водную среду температурой 22–25 оС. Регистрация электрической активности производилась микроэлектродами, заполненными 1 М KCl. Электрическая активность свободно плавающих микроорганизмов (парамеций) регистрировалась в камере объемом 0,35 мл. При помощи микропипетки парамеций переносили в экспериментальную камеру, содержавшую специальный физиологический раствор (KCl – 4 mM, CaCl – 1 mM, MgCl –5 mM, tris HCl – 1 mM. pH раствора – 7,2). Для регистрации применяли стеклянные микроэлектроды, заполненные 0,1 М КСl. Для электрической стимуляции использовали деполяризационные импульсы тока силой 0,1–1 нА, длительностью 1000–3000 мс. Раздражения подавали через второй введенный в микроорганизм микроэлектрод. Так как интактные парамеции чрезвычайно подвижны, регистрация электрической активности с двумя внутриклеточными микроэлектродами (регистрирующим и стимулирующим) была осуществлена всего на трех особях. Микроэлектрод вводили в момент «отдыха» парамеции, чтобы не снижать ее двигательную активность. По этой же причине время регистрации ограничивалось 3–15 мин. Результаты получены на 50 одноклеточных в различных ситуациях поведенческого и электрофизиологического эксперимента. Для опытов с плесневыми грибами применяли бытовую культуру, выросшую в молочнокислой среде. Для эксперимента были специальным образом отобраны образцы поперечного среза, на которых хорошо просматривались слои культуры. Микроэлектроды заполняли 2,5 М KCl. Для работы на нервной системе и полностью изолированных клетках моллюска Helix lucorum применяли стандартную технику микроэлектродной регистрации электрической активности нейронов. Методика подробно описана в ранее опубликованных работах (Греченко, Соколов, 1979), когда в экспериментах была зарегистрирована электрическая активность более 500 нейронов, полностью изолированных или находящихся в составе нервной системы, в различных ситуациях, создаваемых экспериментатором в зависимости от целей исследования, например, при регистрации ритмической активности в течение многих часов при формировании неассоциативных и ассоциативных видов научения.
Результаты
Цианобактерии. Это одноклеточные и нитчатые микроорганизмы-прокариоты, древнейшие представители которых имеют геологический возраст свыше 3,5 млрд. лет. В природе образуют пленки и цианобактериальные маты. При внутриклеточной регистрации электрической активности от индивидуальной цианобактериальной нити обнаружена работа пейсмекерного механизма с регулярными колебаниями, частота которых 0,1–0,5 Гц и амплитудой до 50 мВ (рис. 1, а). Стабильный уровень мембранного потенциала и частота генерации осцилляций сохранялись в течение всего времени наблюдения – более 2 часов. Регистрация электрической активности макроэлектродом в цианобактериальной пленке показывает синхронизированные синусоидальные ритмы частотой около 6 Гц и около 35 Гц. Когда деятельность членов цианобактериального сообщества направлена на решение жизненно важной задачи (например, освоение новых поверхностей, противостояние другому сообществу), их активность синхронизирована, что выражается в генерации высокоамплитудных ритмических колебаний суммарного потенциала (рис. 1, б). При этом за счет движения нитей пленка дифференцирует органоподобные образования. При регистрации потенциалов в зонах, заведомо отличающихся активностью элементов, например, в области формирования тяжей (структурных элементов) и в зонах спокойствия – были получены разные по уровню синхронизированной активности явления.
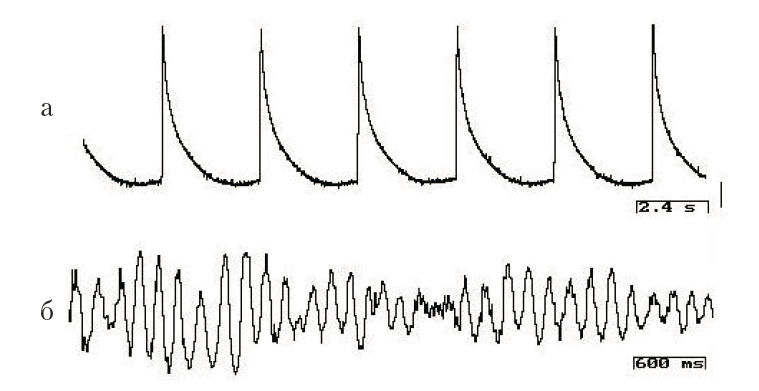
Рис. 1. Электрическая активность цианобактерий Oscillatoria terebriformis, отводимая внутриклеточным (а) и экстраклеточным микроэлектродом (б). Калибровка: 10 мВ, 2,4 с (а), 600 мс (б)
Чем сложнее внутренняя организация живого существа, тем в большей степени метаболические процессы отражаются в эндогенной ритмике: они актуализируются в электрической активности клеток и организуют (или включают) определенные системы исполнения. В наших экспериментах показано, что цианобактерии имеют чрезвычайно устойчивую эндоклеточную электрическую активность, характеризующуюся постоянством периодичности (см. рис. 1, а). Цианобактериальные нити выживают только в сообществе, и синхронизированная электрическая активность является пусковым механизмом совместной деятельности. Как показывают исследования, бактериальные биосоциальные системы характеризуются единым жизненным циклом, следовательно, их активность упорядочить во времени проще, чем, например, у дрожжей. У эволюционно более продвинутых дрожжевых клеток (одноклеточных грибов) внутриклеточная регистрация выявляет наличие сложных паттернов эндогенной активности (см. рис. 2, а, б), среди которых сложно выделить электрические процессы, выражающие циркадианные ритмы.
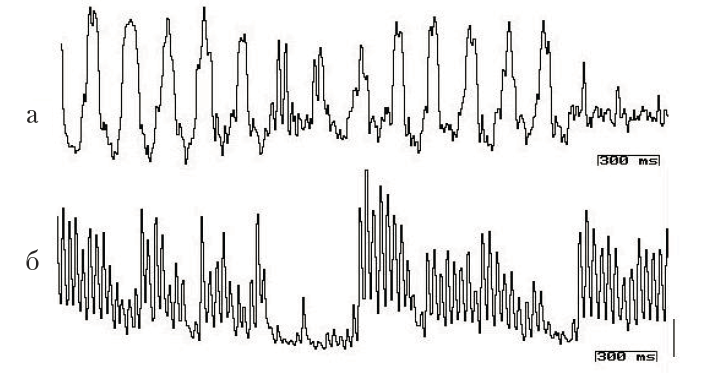
Рис. 2. Электрическая активность дрожжевой клетки Saccharomyces cereviviae, зарегистрированная внутриклеточным микроэлектродом (МЭ), через 3 (а) и 20 (б) мин после начала опыта. Калибровка: 11 мВ, 300 мс
Дрожжи. Одноклеточные эукариоты, грибы. Размеры дрожжевых клеток обычно составляют 3–7 мкм в диаметре. У дрожжей можно выделить высокочастотный компонент до 28 Гц и низкочастотные компоненты около 0,1 Гц (существуют ритмические компоненты с частотой 1/час). В ряде случаев генерация высокочастотных колебаний развивается упорядоченно, группы следуют с частотой 0,8–0,9 Гц. Осцилляции, зарегистрированные внутриклеточным микроэлектродом от индивидуальной дрожжевой клетки, характеризуются чрезвычайным разнообразием как по амплитуде, так и по форме и временной организации (рис. 2). Паттерны активности повторяются с определенной частотой – каждый вид активности отличается индивидуальной периодичностью. На электрические процессы дрожжевых клеток влияют такие факторы внешней среды, как температура, состав жидкостной среды в экспериментальной камере и длительность нахождения в растворе определенного состава. В результате проведенных опытов показано, что величина заряда клетки изменяется в зависимости от цикла активности, осуществляемого организмом.
Парамеции. Это свободноживущие одноклеточные эукариоты, наиболее древние представители которых появились примерно 2,5 млрд. лет назад. Клетки имеют форму туфельки длиной 0,12–0,13 мм. Исследователями накоплен громадный материал о структуре различных клеточных элементов, свойствах электровозбудимой цитоплазматической мембраны, белковом составе и свойствах отдельных белков, входящих в систему управления двигательной активностью парамеций.
Результаты регистрации электрической активности Paramecium caudatum внутриклеточным микроэлектродом указывают на то, что уровень мембранного потенциала покоя (ПП) этих существ характеризуется значительными колебаниями: обнаружены изменения от –40–45 мВ до +20 мВ, периоды стабильного значения ПП чрезвычайно короткие – в условиях проведенных опытов не более 30–70 с. Анализ электрической активности показывает, что, во-первых, в клетке регистрируются медленные и быстрые осцилляции, во-вторых, одновременной активностью, как правило, обладают несколько эндогенных осцилляторов; эти данные согласуются с наблюдениями других исследователей, экспериментировавших на этом объекте (Доронин, Зазулин, 1976). На некоторых записях можно выделить два-три одновременно работающих пейсмекера ( рис. 3, а, б). Увеличение амплитуды волн эндогенной активности может приводить к генерации потенциалов действия (ПД) разной амплитуды (см. рис. 3, а). Амплитуда ПД, по-видимому, зависит от функциональных особенностей вызвавшего его пейсмекера. На нейрограммах можно видеть ПД как одинаковой, так и различной амплитуды, отличающиеся единицами и даже десятками мВ (рис. 3, а–в). Частота зарегистрированных колебаний от 0,1кол/сек до 20 кол/сек, амплиту
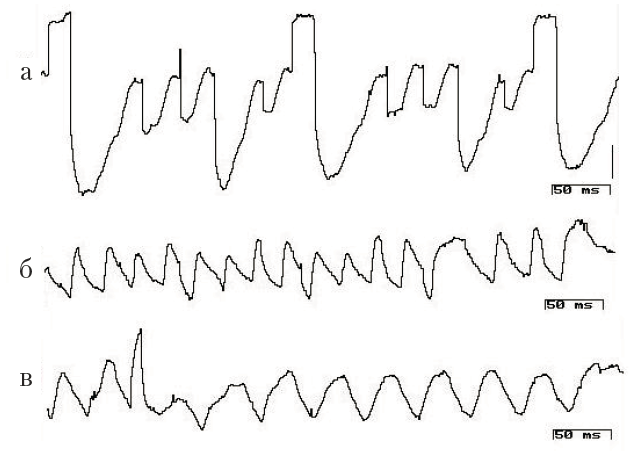
Рис. 3. Электрическая активность парамеции, зарегистрированная в одном опыте. Интервалы между регистрациями 5–7 мин. Калибровка: 11 мВ, 50 мс
Плесневые грибы. В отличие от бактерий и одноклеточных эукариот, плесень состоит из множества клеток, которые образуют длинные нити (гифы). Под микроскопом плесень выглядит как сплетение разветвленных гиф, образующих мицелий (рис. 4, д). Часть гиф оканчивается спорами, они определяют цвет плесени. У плесени имеются «ветви» и «корни», похожие на тонкие нити. Отростки плесневых грибов Penicillium образуют слои, каждый из которых характеризуется особым видом электрической активности, отличающимся паттерном осцилляций, частотой их генерации и амплитудой (см. рис. 4). При исследовании плесневых грибов, образующих своими отростками сложноорганизованные системы, обнаружены чрезвычайно многообразные формы электрической активности (см. рис. 4, а).
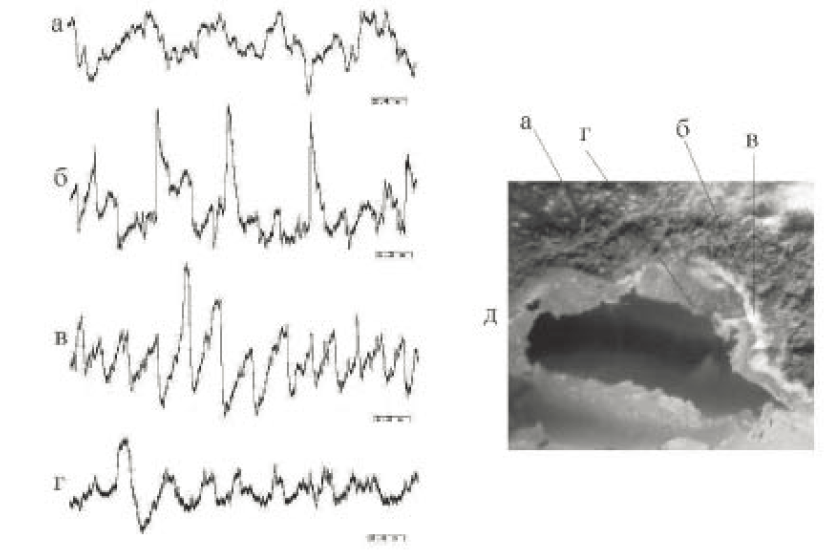
Рис. 4. Электрическая активность, зарегистрированная в разных слоях плесневого гриба: а–г – электрическая активность в слоях (д) соответственно; д – фотография слоев, в которых производилась регистрация. Калибровка: 10 мВ, 2,4 с (а, г), 1,2 с (б, в)
Пейсмекеры моллюсков. О пейсмекерной активности нейронов моллюсков известно чрезвычайно много, так как именно у улиток были впервые обнаружены ритмически генерирующие клетки (Arvanitaki, Chalazonitis, 1964). Одна из первых работ, в которой был зарегистрирован циркадианный ритм пейсмекерного нейрона, выполнена Штрумвассером и Уилсоном на препарате моллюска аплизии (Strumwasser, Wilson, 1976). При регистрации электрической активности в изолированном париетовисцеральном ганглии было показано, что идентифицированный нейрон R15 имеет потенциалы, ритмичность которых зависит от цикличности света и темноты.
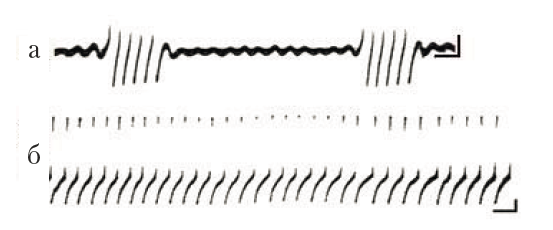
Рис. 5. Пейсмекерная активность изолированных нейронов наземного моллюска Helix lucorum: групповая (а) и регулярная (б) активность, зарегистрированная внутриклеточным МЭ. Калибровка: 10 мВ, 1 с
Наши опыты показали, что пейсмекерная электрическая активность характерна для многих нейронов ЦНС моллюсков, а функциональное разнообразие задач, в которые вовлекаются такие нейроны, исключительно велико (Греченко, 2008). В частности, клетки пейсмекерного типа участвуют не только в регуляции функциональных состояний, зависящих от времени суток, но и в осуществлении различных движений в качестве командных единиц.
Обсуждение
Все исследованные нами организмы имеют осцилляторную эндогенную активность, которая проявляется в виде циклических изменений мембранного потенциала. Какова функциональная роль этих осцилляций, являются ли они электрофизиологическим эквивалентом биологических часов? Однозначный ответ на этот вопрос дать сложно, так как длительность регистрации электрической активности была слишком мала, однако и исключить такую возможность нельзя (см. рис. 1). Значение регулярных колебаний мембранного потенциала, по-видимому, различно: пейсмекерные волны отражают течение метаболических процессов, которые не обязательно связаны с генами биологических часов. Например, изучая методами флюоресцентной микроскопии развитие биохимических реакций у дрожжевых клеток, исследователи выделили несколько циклических превращений (Richard, 2003). Хотя дрожжи – одноклеточные и сравнительно простые организмы, они имеют чувство времени, которое не связано с репродуктивными циклами. Гликолитический путь (десять последовательных реакций, каждая из которых катализируется отдельным ферментом) демонстрирует ритмичность процессов. Данный факт означает, что метаболические концентрации осциллируют вокруг фосфофруктокиназы – одного из важнейших ферментов гликолиза в тканях организма. Частота этих осцилляций – около минуты, если используются интактные клетки. Экстракт дрожжевых клеток тоже осциллирует, хотя и с меньшей частотой. У интактных клеток также наблюдаются макроскопические ритмические колебания при одновременном совместном осциллировании большинства клеток. Длительные осцилляции требуют работы синхронизирующего механизма, который вовлекает ацетальдегид в качестве сигнального компонента, действующего в определенных условиях.
Очевидно, все существа, независимо от их организации и положения в системе живых организмов, обладают памятью: так, например, было обнаружено существование двух видов памяти у растений (Thellier and Luttge, 2013).
Анализ информационного содержания памяти выделяет ее процедурную и декларативную составляющие. С этой позиции многие жизненные функции любого живого существа можно рассматривать как проявление процедурной памяти – пластические изменения нейронных ответов при обучении любого вида и сами нейронные ответы, различные вариации поведения в определенных условиях, зависимость состояния организма от времени суток, реализация его возможностей и пр. Благодаря наличию и работе древней памяти стремление к выживанию стало обладать фундаментальной связью между воздействием и адекватным ответом организма. Возникновение и сохранение жизни основано на формировании и развитии этой «базисной» памяти, а совершенствование живых существ происходит благодаря наращиванию её уровней (памяти выживания). Все проявления памяти имеют молекулярную основу, на этом базируются и рецепция сигналов, и их передача, запоминание и дальнейшее применение распознанной информации. Хранение информации и ее узнавание осуществляется сложными ансамблями элементов с высокой степенью интеграции и обратной связью, формирующими «системный» уровень организации памяти. Однако основу памяти составляют процессы, происходящие во внутренних пространствах органических компонентов, из которых строится живое существо: кальциевые волны, эпигенетические модификации ДНК и гистонов, процессы регуляции различных согласований (расписания синхронизаций) через биологические часы (Gerstner, 2012). Изучение синхронизации генетических или клеточных осцилляторов является центральной проблемой понимания ритмичности живых организмов на молекулярном и клеточном уровне. В исследованиях синхронизации генетических осцилляторов (Zhou et al., 2008) показано, каким образом формируется коллективный ритм в популяции генетических осцилляторов через синхронизирующие внутриклеточные коммуникации и каким образом ансамбль независимых генетических осцилляторов синхронизируется посредством шумящих сигнальных молекул. Основная задача исследований состоит в том, чтобы показать роль синхронизирующих внутриклеточных механизмов с точки зрения их динамических свойств на основании изучения коммуникаций различных органических компонентов, нескольких видов шума и внешних стимулов. Стохастические стимуляции показывают, что устойчивые циркадианные осцилляции могут возникать на клеточном уровне даже в том случае, когда максимальное число мРНК и молекул протеинов, вовлекаемых в осцилляции, не превышает порядка нескольких десятков или сотен (Gonze et al., 2003).
Среди множества ритмов, генерируемых клетками, биологические часы являются наиболее изученными. Многие примеры клеточных циркадианных систем подтверждают, что часы – это не просто эмерджентное качество сложной системы, действие данного механизма основано на особых свойствах индивидуальных клеток. Предположение о генетической основе свободнотекущих циркадианных ритмов возникло еще в 1932 году (Bunning, 1932). Результаты исследований мутантов Drosophila melanogaster позволили идентифицировать часовой ген (Konopka, Benzer, 1971). С тех пор были обнаружены многие часовые гены у организмов разного уровня эволюции (Merrow et al., 2005; McClung, 2006). У большинства млекопитающих ряд часовых генов и протеинов формируют регуляторную систему, которая продуцирует осцилляции циркадианного периода – 24 часа. Молекулярные и физиологические ритмы координируются с суточными изменениями окружающей среды посредством доминирующего циркадианного пейсмекера супрахиазменного ядра гипоталамуса. Нейроны этой структуры эндогенно генерируют циркадианный ритм и приспосабливают его к смене циклов день-ночь окружающей среды. Стремление понять, как такая гетерогенная сеть продуцирует когерентный синхронный циркадный выход, является мотивом проведения многочисленных экспериментальных и теоретических работ. Хотя все циркадианные системы составлены из клеточных часов, в их организации существуют большие различия. Например, у растений циркадианные ритмы соседних клеток, по-видимому, идут в независимых фазах, а у животных они формируются на основе внутренне связанных иерархических систем. Новые данные открывают возможность для предположений, что молекулярный механизм циркадных часов у цианобактерии Synechococcus elongatus составлен из множества осцилляторных систем (как это было описано для моделей часовых механизмов эукариот). Но альтернативная интерпретация состоит в том, что работа пейсмекерного механизма, как это предполагалось ранее, основывается и зависит от скорости гидролиза АТФ посредством часового протеина KaiC (Brunner et al., 2008). Цианобактериальные часы имеют все те же самые фундаментальные свойства, что и эукариотические часы, однако их компоненты не полностью гомологичны тем, которые обеспечивают измерение времени у животных, растений и грибов (Eelderink-Chen et al., 2010; Merrow, Raven, 2010).
Мы полагаем, что полученные нами результаты могут свидетельствовать об исходном наличии жесткой зависимости между внутриклеточными процессами, инициированными некоторыми «ключевыми» факторами внешней и внутренней среды и действиями организма. Это – азбука существования, заложенная в самый фундамент развития живого вещества: без такой согласованности внешних и внутренних процессов организм не выживает.