Задача исследования - изучение психофизиологических процессов, которые могут вызывать физиологический и временной эффект Струпа при наличии и отсутствии реакции подавления в ответ на предъявление одного из стимулов. Участники исследования (n = 26) выполняли две задачи Струпа, стимулами для которых служили слова «зеленый» или «красный», написанные зеленым или красным шрифтом. В задаче 1 нужно было опознавать стимулы, нажимая разные кнопки в зависимости от цвета шрифта. В задаче 2 нужно было игнорировать слово «красный», написанное красным или зеленым шрифтом; слово «зеленый» также распознавалось по цвету шрифта. Регистрировались время реакции и вызванные потенциалы на стимулы. В результате обнаружен временной эффект Струпа в ответ на предъявление слова «красный» (в задаче 1), в ответ на предъявление слова «зеленый» (в задаче 2). Время реакции в ответ на предъявление слова «зеленый» в задаче 2 на 113 мс больше, чем в задаче 1. Различия в амплитуде компонентов вызванных потенциалов P50, N100, P300 в ответ на предъявление данных стимулов позволяют локализовать эффект Струпа на ранней и поздней латентности и свидетельствуют о различии в психофизиологических процессах, которые участвуют в выполнении задачи при наличии и отсутствии реакции подавления ответа.
В 1935 г. Дж. Струп обнаружил, что время называния цвета графических стимулов (квадратов) оказывается меньшим, чем время называния цвета шрифта неконгруэнтных словесных стимулов, обозначающих цвета, не совпадающие с цветом написанных слов (Stroop, 1935). Разность времени называния цвета шрифта неконгруэнтных слов и времени называния цвета окрашенных квадратов составляет классический эффект Струпа.
В нашей работе изучался модифицированный временной эффект Струпа, величина которого определялась разностью во времени между распознаванием цвета (двигательный ответ) двух неконгруэнтных слов - «красный» и «зеленый». Ранее было установлено, что коэффициент корреляции между классическим и модифицированным эффектом Струпа при назывании цветов этих слов достаточно большой (г = 0,84) и значим при р < 0,01 (Беспалов, Климова, 2014).
Существует также физиологический эффект Струпа, проявляющийся в различиях вызванных потенциалов (далее - ВП) при распознавании по цвету шрифта конгруэнтных и неконгруэнтных стимулов (Беспалов, Вартанов, 2014) Данный эффект может быть обусловлен процессом обнаружения (detection) испытуемыми противоречия или конфликта между значением слова и цветом его шрифта, находящего отражение в негативной волне ВП на латентности 350-500 мс (Badzakova-Trajkov et al., 2009; Liotti et al., 2000). Отмечено возрастание амплитуды негативного компонента N450 в неконгруэнтном условии, где увеличивается конфликт между значением и цветом слова (Ergen et al., 2014).
Некоторые авторы соотносят физиологический эффект Струпа с артикуляторной и фонологической обработкой стимула на латентности 300-450 мс (Zurron et al., 2009); другие обнаруживают позитивный компонент Р300 на латентности 300-450 мс, негативно коррелирующий со сложностью задачи и уменьшающийся на неконгруэнтные стимулы по сравнению с конгруэнтными (Houston, Bauer, Hesselbrock, 2004; Potter et al., 2002). Этот компонент соотносится с переходом от восприятия конфликта к принятию решения об ответе.
Известно, что задача Струпа используется при исследовании когнитивного контроля - метакогнитивного процесса, который инициирует, управляет и координирует другие когнитивные процессы, направленные на осуществление целенаправленного поведения (Величковский, 2009; Alvarez, Emory, 2006). Основные функции когнитивного контроля (в соответствии с моделью Мияке) - переключение между задачами, подавление, обновление информации (Miyake et al., 2000). С активацией произвольного подавления при выполнении задачи Струпа могут быть связаны изменения ВП на латентности 200-400 мс (Liotti et al., 2000). Фактор «подавление ответа» исследуется также в экспериментальной парадигме «Go/NoGo»: требуется реагировать на один стимул, но подавлять ответ на другой; частный случай «Go/NoGo» - задача «Стоп-сигнал» (Logan, 1994). Исследований с совместным использованием парадигмы «Стоп-сигнал» и задачи Струпа представлено достаточно мало. Так, обнаружено увеличение среднего времени реакции в задаче Струпа, осложненной «Стоп-сигналом» (Verbruggen, Liefooghe, Vandierendonck, 2004). Сочетание разных парадигм позволяет более детально изучить психофизиологические процессы, вовлеченные в процесс их выполнения.
В нашем исследовании мы скомбинировали парадигмы Струпа и «Стоп-сигнал». Одна экспериментальная задача решалась испытуемыми без подавления ответа на стимулы Струпа, а в другой задаче требовалось подавлять ответ на некоторые стимулы. Основная задача исследования состояла в изучении психофизиологических процессов, которые могут вызывать физиологический и временной эффект Струпа, регистрируемый при наличии и отсутствии подавления на один из стимулов. Эмпирическое изучение этих процессов проводилось с помощью регистрации и анализа отношений между показателями времени реакции (далее - ВР) и ВП в разных экспериментальных задачах. Изучался вопрос о том, всегда ли влияние на ВР фактора «конгруэнтность стимулов» (т. е. временной эффект Струпа) должно сопровождаться физиологическим эффектом Струпа в определенных отведениях и компонентах ВП, и наоборот.
Теоретическое объяснение полученных экспериментальных данных осуществлялось нами с помощью сферической нейросетевой модели опознавательного действия, в которой выделен и описан ряд психофизиологических процессов, связанных с разными нейронными ансамблями мозга (Беспалов, 2014). Эта модель основана на идее «векторного кодирования» в мозге информации о тестовых стимулах и ответов на них (Соколов, Вайткявичус, 1989; Фомин, Соколов, Вайткявичус, 1979). Предполагается, что состав и последовательность включенных в опознавательное действие нейронных ансамблей и соответствующих им психологических процессов определяется задачей испытуемого и условиями опознания, при варьировании которых могут изменяться показатели ВР и ВП. В разных задачах и условиях опознавательное действие осуществляется с помощью разных психофизиологических процессов. Если теоретическая реконструкция этих процессов, осуществляемая с позиции сферической нейросетевой модели, позволит раскрыть возможные причины и механизмы возникновения наблюдаемых особенностей показателей ВР и ВП, это может свидетельствовать об адекватности этой модели и «аккумулированных» в ней идей Е.Н. Соколова и др.. эмпирическая проверка которых также составляла одну из задач данной работы.
Методика
Испытуемые. В исследовании приняли участие 26 испытуемых мужского пола с высшим или неоконченным высшим образованием, для которых русский язык являлся родным. Средний возраст участников составил 23,3 ± 3,7 лет. Все испытуемые имели нормальное или скорректированное до нормального зрение.
Аппаратура. Программирование экспериментальных заданий и предъявление стимулов и регистрация ВР на них осуществлялись с помощью программы Presentation. Для показа стимулов использовался профессиональный монитор с разрешением 800 х 600 пикселей и частотой вертикальной развертки 200 Гц, расположенный на расстоянии 40-45 см от испытуемого. Для регистрации ВР использовалась лазерная двухкнопочная USB-мышь (Razer Abyssus Black), частота опроса которой составляла 1000 Гц. Для регистрации ВП ЭЭГ использовался энцефалограф «Нейро-КМ» фирмы «Статокин» (19 каналов, система 10-20%, референтные электроды - мастоиды М1 и М2, заземляющий электрод на Fpz, сопротивление 3-30 кОм).
Схема эксперимента. Стимулами Струпа были 4 слова: «красный» и «зеленый», написанные красным или зеленым шрифтом. В двух конгруэнтных стимулах цвет шрифта слова согласуется с его семантикой (т. е. обозначаемый словом цвет совпадает с цветом шрифта, которым написано слово), а в неконгруэнтных (конфликтных) стимулах цвет и семантика слов различны. Стимулы предъявлялись в квазислучайном порядке по одному в центре монитора на 1,2 с. Временной интервал между выключением одного стимула и началом предъявления другого составлял 1 с, так что одна проба занимала 2,2 с.
Каждый испытуемый решал задачи 1 и 2. В задаче 1 испытуемый опознавал каждый стимул Струпа по цвету шрифта. В ответ на предъявление стимулов с зеленым цветом шрифта испытуемый должен был нажать левую кнопку мыши, а в ответ на предъявление красных по цвету слов испытуемый должен был нажать правую кнопку. В задаче 2 испытуемый должен был игнорировать слово «красный», написанное красным или зеленым шрифтом, не нажимая в ответ на его предъявление кнопку мыши. Однако «зеленые» по семантике слова он должен был опознавать по цвету шрифта, как и в первой задаче (в ответ на предъявление зеленого по цвету шрифта слова «зеленый» он должен был нажать левую кнопку мыши, а в ответ на предъявление красного по цвету слова «зеленый» - правую кнопку). В одной задаче каждый стимул Струпа предъявлялся по 70 раз. Одна задача содержала 280 проб и длилась около 10 мин.
Регистрируемые показатели и их обработка. В ходе эксперимента фиксировалось время выполнения задач - ВР, а также показатели ЭЭГ- ВП, возникающие в ответ на предъявление каждого стимула. Полученные ВП обрабатывались с помощью программы «BrainSys». Для анализа ВП был взят отрезок записи, начиная с 200 мс до стимула (появление слова) и заканчивая 500 мс после показа стимула. Была проведена сортировка выделенных отрезков ЭЭГ по номерам стимулов и удалены фрагменты, содержащие артефакты. Для каждого участника в каждой задаче Струпа записи ВП усреднялись по каждому из четырех стимулов и в каждом из 19 отведений. Затем эти индивидуальные ВП усреднялись (с оценкой значимости различий) по группе участников по четырем стимулам и для каждой задачи в отдельности. Оценка значимости различий между ВП на стимулы в разные моменты времени, а также различий между ВР на разные стимулы и в разных задачах проводилась с помощью программы «Statistica» и t-критерия Стьюдента.
Зависимыми переменными были регистрируемые показатели ВР и ВП; в качестве независимых переменных выступали два двухуровневых фактора: а) фактор «конгруэнтность стимула», связанный с совпадением (конгруэнтность) или несовпадением (неконгруэнт- ность) семантики и цвета шрифта стимула; б) фактор «подавление ответа», связанный с отсутствием (задача 1) или наличием (задача 2) необходимости подавлять ответ на слово «красный», написанное красным или зеленым шрифтом.
Результаты
Времена реакции (ВР). Средние групповые ВР на тестовые стимулы в задачах 1 и 2 показаны в табл. 1. В задаче 1 средний по группе временной эффект Струпа на слове «красный» равен 24 мс и значим при р < 0,001 по критерию Стьюдента. Аналогичный эффект для слова «зеленый» в задаче 1 равен -4 мс и незначим (р = 0,43). При этом средние по двум стимулам ВР на слово «зеленый» (451 мс) и на слово «красный» (447 мс) не различаются по t-критерию для зависимых выборок (р = 0,146). В задаче 2 эффект Струпа на слове «зеленый» увеличился до 15 мс и стал значимым при р = 0,006 по критерию Стьюдента. Кроме того, в задаче 2 среднее ВР на слово «зеленый» равно 564 мс и на 113 мс больше (при р < 0,001), чем ВР на аналогичное слово в задаче 1 (451 мс).
Вызванные потенциалы (ВП). В табл. 2 и 3 звездочками (*) отмечены различия между амплитудами ВП, значимые по критерию Стьюдента на некоторых латентностях и вызываемые изучаемыми в работе факторами. Оказалось, что такие различия имеются на трех латентностях - около 50 мс, 100 мс и 250-500 мс. Таким образом, анализировались следующие компоненты вызванных потенциалов: Р50 (латентность примерно 50 мс после возникновения стимула), N100 (латентность 80-120 мс), Р300 (латентность 250-500 мс) (Polich, 2007).
Влияние на ВП фактора «подавление ответа». Различия между компонентами ВП на тестовые стимулы в задаче 1 и 2 оказались значимыми (при p < 0,05) на разных латентностях (табл. 2). Эти различия можно трактовать как физиологические эффекты фактора
Таблица 1
Средние групповые ВР (мс) на тестовые стимулы в задачах 1 и 2
|
Задача 1 (отсутствует подавление ответа на стимулы) |
Задача 2 (возникает подавление ответа на слово «красный») |
|||||||
|
Конгруэнтные стимулы |
Неконгруэнтные стимулы |
Временной эффект Струпа на слове «красный» |
Временной эффект Струпа на слове «зеленый» |
Конгруэнтный стимул |
Неконгруэнтный стимул |
Временной эффект Струпа на слове «зеленый» |
||
|
Слово «красный» красным шрифтом |
Слово«зеленый» зеленым шрифтом |
Слово «красный» зеленым шрифтом |
Слово «зеленый» красным шрифтом |
Слово «зеленый» зеленым шрифтом |
Слово «зеленый» красным шрифтом |
|||
|
435 |
453 |
459 |
449 |
24 р < 0,001 |
-4 р = 0,146 |
557 |
572 |
15 р = 0,006 |
«подавление ответа». Из табл. 2 видно, что для конгруэнтного слова «красный» различия в амплитуде компонента P50 значимы в четырех отведениях (О2, Т4, Т6 и Р4), тогда как для неконгруэнтного слова «красный» различия в этом компоненте ВП наблюдаются только в отведении О2. Для слова «зеленый» значимых различий между задачами в амплитуде компонента P50 не обнаружено. Для компонента P300 различия между задачами в табл. 2 приведены только для тех отведений, в которых они значимы для обоих тестовых слов.
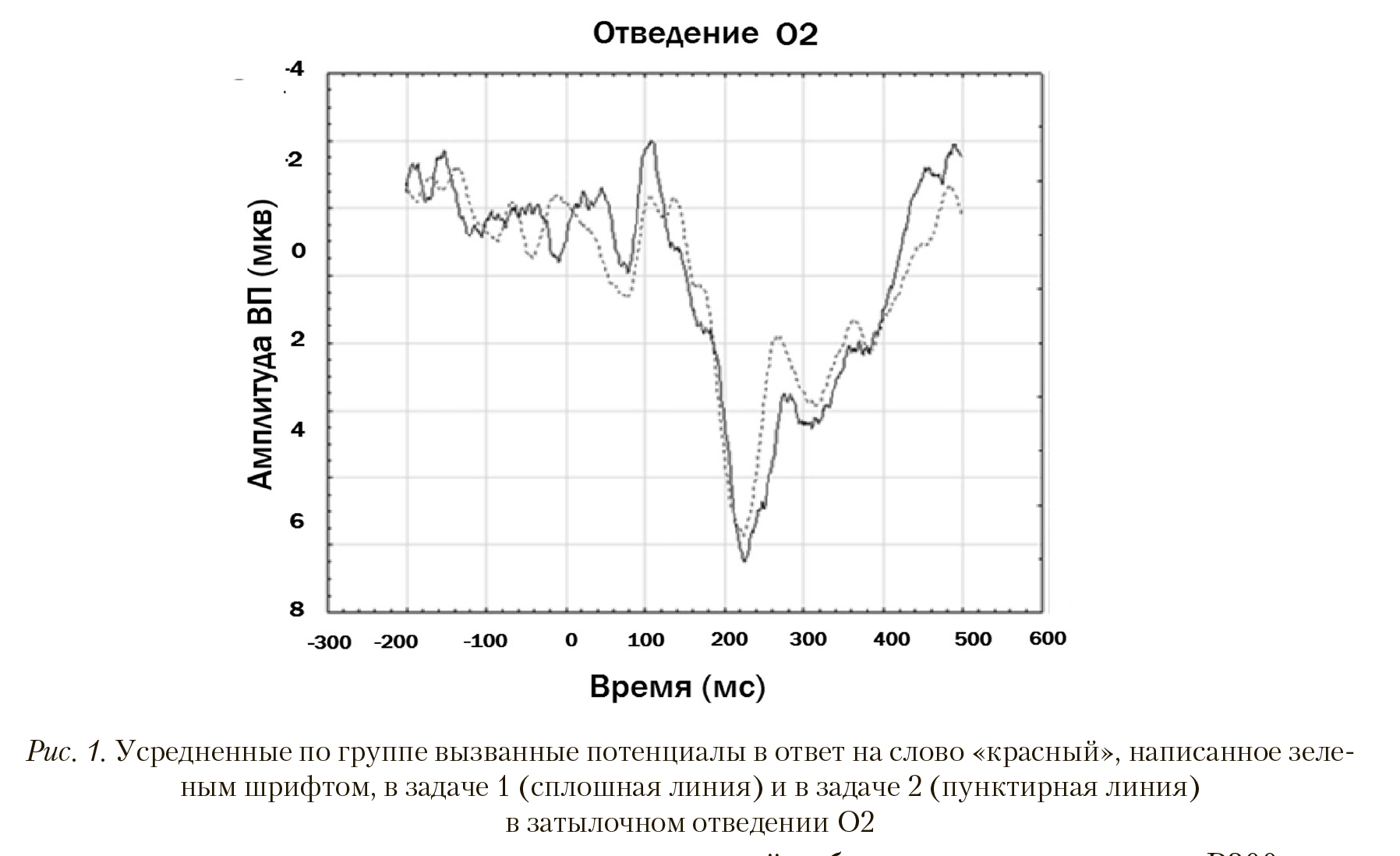
Графики ВП, например, в отведении О2 для задач 1 и 2 на слово «красный», написанное зеленым шрифтом, показаны на рис. 1. Несмотря на то, что на латентностях 100, 270 и 450 мс эти графики визуально сильно различаются, статистически значимое различие (отраженное в табл. 2) имеется только на латентности 56-59 мс, или в позитивном компоненте Р50.
Еще один пример графиков ВП в другом отведении F3 для задач 1 и 2 и слова «зеленый», написанного красным шрифтом, показан на рис. 2. Статистически значимое различие этих графиков имеется на латентности 401-448 мс, т. е. в компоненте Р300, который относится к диапазону от 250 до 500 мс.
Влияние на ВП фактора «конгруэнтность стимула» (физиологический эффект Струпа). Значимое различие между ВП (при p < 0,05 по критерию Стьюдента) в задаче 1
Таблица 2
Значимые (*) при p < 0,05 по критерию Стьюдента различия в амплитуде вызванных потенциалов в задачах 1 и 2, обусловленные фактором «подавление ответа»
|
Компонент |
Отведение |
Тестовые стимулы |
|||
|
Слово «зеленый» зеленым шрифтом |
Слово «зеленый» красным шрифтом |
Слово «красный» зеленым шрифтом |
Слово «красный» красным шрифтом |
||
|
P50 |
02 (правое затылочное) |
Нет |
Нет |
* |
* |
|
Т4 (правое височное) |
Нет |
Нет |
Нет |
* |
|
|
Т6(правое височное) |
Нет |
Нет |
Нет |
* |
|
|
Р4(правое теменное) |
Нет |
Нет |
Нет |
* |
|
|
N100 |
02 (правое затылочное) |
* |
Нет |
Нет |
* |
|
Т4 (правое височное) |
Нет |
* |
Нет |
Нет |
|
|
Г/ (фронто-центральное) |
Нет |
* |
Нет |
Нет |
|
|
P300 |
Р4(правое теменное) |
* |
* |
* |
* |
|
Т4 (правое височное) |
* |
* |
* |
* |
|
|
Т5 (левое височное) |
* |
* |
* |
* |
|
|
Г8 (правое фронтальное) |
* |
* |
* |
* |
|
|
Т6(правое височное) |
* |
Нет |
* |
* |
|
на конгруэнтное и неконгруэнтное слово «красный» обнаружено в компоненте Р300 в отведении Т3 (табл. 3). Этот результат согласуется с данными исследований, свидетельствующими о том, что физиологический эффект Струпа также локализуется в компоненте P300 (Houston, Bauer, Hesselbrock, 2004; Potter et al., 2002). При этом в задаче 2 различие ВП в
ответ на предъявление конгруэнтного и неконгруэнтного слова «красный» обнаружено в другом компоненте Р50 в отведении Т6. Вместе с тем, значимые различия между ВП в ответ на предъявление конгруэнтного и неконгруэнтного слова «зеленый» в задаче 1 отсутствуют на всех латентностях (табл. 3). Этот результат согласуется с отсутствием в данной задаче временного эффекта Струпа на данное слово. В задаче 2 значимые различия между ВП в ответ на предъявление конгруэнтного и неконгруэнтного слова «зеленый» обнаружено только для компонента Р50 в отведении О2. В этой задаче имеется также временной эффект Струпа (табл. 1).
Таблица 3
Значимые (*) по критерию Стьюдента различия (при p < 0,05) в амплитуде
компонентов вызванных потенциалов, обусловленные фактором
«конгруэнтность-неконгруэнтность стимулов»
|
Слово |
Отведение |
Компонент ВП |
Задача 1 |
Задача 2 |
|
«Красный» |
Т6(правое височное) |
P50 |
Нет |
* |
|
Т3 (левое височное) |
P300 |
* |
Нет |
|
|
«Зеленый» |
О2 (правое затылочное) |
P50 |
Нет |
* |
Обсуждение результатов
Влияние на ВР и ВП фактора «подавление ответа». Для объяснения того факта, что в задаче 2 среднее ВР на слово «зеленый» увеличилось на 113 мс по сравнению с задачей 1, можно предположить, что в задаче 2 в опознавательное действие включен дополнительный процесс, состоящий в идентификации (оценке) тестовых стимулов как слов «зеленый» или «красный», с которыми в данной задаче с помощью инструкции соотнесены разные последующие процессы. Если стимул идентифицируется как слово «зеленый», то вслед за этим должен осуществляться процесс определения цвета его шрифта. Если же стимул идентифицируется как слово «красный», то должен осуществляться процесс отказа от двигательного ответа. Эти процессы требуют дополнительной активности и времени, что приводит к увеличению ВР на слово «зеленый» в задаче 2. (В задаче 1 для определения цвета шрифта стимула не требуется его предварительная идентификация как слова «зеленый» или «красный».)
Можно предположить, что предварительная идентификация стимулов в задаче 2 (где требуется подавление ответа) может осуществляться на основе определения их начертания, или выделения графической формы. Этот процесс должен проявляться в изменениях ВП на ранней латентности. Такие различия ВП действительно наблюдаются. Так, в задачах 1 и 2 ВП на слово «красный» различаются на латентности около 50 мс в правом затылочном отведении О2 и др. (табл. 2), а ВП на слово «зеленый» различаются на латентности около 100 мс в правом височном Т4 и в фронто-центральном отведении Fz (табл. 2).
В идентификации тестовых стимулов в задаче 2 может принимать участие также внутренняя артикуляция стимулов, которая выполняется после или одновременно с выделением их графической формы. Поскольку этот процесс отсутствует в задаче 1 (где отсутствует необходимость в предварительной идентификации стимулов), то можно ожидать, что ВП на одинаковые слова (зеленый-зеленый и красный-красный) в задачах 1 и 2 будут различаться на латентности около 100 мс. Это действительно наблюдается в трех отведениях - O2, T4 и Fz (табл. 2).
Таким образом, различия между ВП в задачах 1 и 2 на латентностях 50 и 100 мс, а также различия между ВР в этих задачах обусловлены необходимостью подавлять двигательный ответ в задаче 2, что порождает включение в эту задачу дополнительного процесса предварительной идентификации стимула. Этот процесс осуществляется с помощью двух актов: а) выделение графической формы стимула, которое проявляется в раннем позитивном пике (Р50), и б) процесс внутренней артикуляции тестовых стимулов, который проявляется в более позднем негативном пике N100.
На поздних латентностях (в компоненте Р.300) различия между ВП в задачах 1 и 2 наблюдаются для всех тестовых стимулов в четырех отведениях (Р4, Т4, Т5 и F8). Это свидетельствует о том, что в этих задачах тестовые стимулы на латентности после 250 мс обрабатываются с помощью разных психофизиологических процессов, связанных с нисоншими, теменными и фронтальными областями мозга. Например, различие процессов обработки слова «красный» на латентности после 250 мс в задачах 1 и 2 может состоять в том, что в эти моменты времени в задаче 1 завершается процесс определения цвета шрифта данного слова и начинается процесс организации двигательного ответа. Сам же двигательный ответ в этой задаче дается в среднем через 447 мс. Вместе с тем, в задаче 2 двигательный ответ на слово «красный» свсдндляетть Поэтому в задаче 2 на латентности более 250 мс процессы определения цвета шрифта слова «красный» и организации ответа на это слово не выполняются. На физиологическом уровне это проявляется в значительных различиях между ВП на этой латентности в задачах 1 и 2.
Процессы обработки слова «зеленый» на латентности более 250 мс в задачах 1 и 2 также различны. Если в задаче 1 в некоторый момент времени завершается процесс определения цвета шрифта слова «зеленый» и начинается процесс организации двигательного ответа, то в задаче 2 эти же процессы должны выполняться примерно на 120 мс позже, поскольку двигательный ответ на слово «зеленый» в задаче 2 дается на 120 мс позже, чем в задаче 1. Значительные различия в психофизиологических процессах, выполняемых в задачах 1 и 2 на латентности 250-500 мс, обусловливают значимые различия ВП, регистрируемые для всех стимулов в большом числе отведений на данной латентности (табл. 2).
Дополнительное объяснение тому факту, что ВР в ответ на предъявление слова «зеленый» в задаче 2 увеличивается на 113 мс по сравнению с задачей 1, связано с изменением вероятности предъявления «целевых» символов. «Целевые» стимулы требуют двигательного ответа (нажатия кнопки мыши), а «нецелевые» игнорируются (Величковский и др., 2016) и не требуют двигательного ответа. В задаче 1 все стимулы являются «целевыми», тогда как в задаче 2 (осложненной «Стоп-сигналом») половина стимулов (слово «красный») не являются «целевыми». Таким образом, «целевые» стимулы предъявляются в задаче 2 в 50% случаев (слово «зеленый»). При предъявлении таких стимулов нужно правильно соотнести цвет слова с заданной кнопкой ответа. Более редкие «целевые» стимулы могут вызывать увеличение ВР и изменение характеристик ВП. Увеличение амплитуды ВП при появлении редких стимулов по сравнению с частыми показано в других работах (Duncan-Johnson, Donchin, 1977; Squires, Squires, Hillyard, 1975; van den Wildenberg, van Boxtel, van der Molen, 2003).
Произвольное подавление ответа рассматривается также как один из механизмов осуществления когнитивного контроля (Величковский и др., 2016). Произвольное подавление требует увеличения ресурсов внимания к более редким «целевым» стимулам в задаче 2, так как необходимо постоянно переключаться между необходимостью подавлять двигательный ответ в ответ на предъявление слова «красный» и давать такой ответ в ответ на предъявление слова «зеленый». С фактором подавления ответа связаны различия ВП на поздней латентности (компонент Р300), регистрируемые для всех стимулов в большом числе отведений (табл. 2).
В то же время, несмотря на подавление двигательного ответа на «нецелевые» стимулы в задаче 2, цвет их шрифта, вероятно, все же выделяется, но на неосознаваемом, автоматическом уровне, о чем могут свидетельствовать указанные в табл. 3 различия ВП между конгруэнтным и неконгруэнтным словами «красный» в задаче 2 на ранней латентности. Такая цветовая обработка происходит в правом полушарии, о чем свидетельствуют различия между ВП в правом височном отведении T6.
Влияние на ВР и ВП фактора «конгруэнтность стимулов». В задаче 1 наблюдается значимый временной эффект Струпа в ответ на предъявление слова «красный», который сопровождается физиологическим эффектом Струпа в левом височном отведении Т3 в компоненте Р300 (на латентностях 370-375, 397-413, 440-444 мс). Этот результат может свидетельствовать о том, что временной эффект Струпа, или различия ВР в ответ на предъявление неконгруэнтного и конгруэнтного слова «красный», связан с различиями процессов обработки данных стимулов в височных областях мозга, происходящими после 370 мс после показа слова «красный». Таким процессом может быть внутренняя артикуляция цвета шрифта тестового слова для его последующего соотнесения с опознавательными эталонами. Поскольку процесс артикуляции цвета шрифта неконгруэнтного (конфликтного) слова «красный» выполняется на фоне непроизвольного внутреннего чтения этого слова и эти артикуляции не совпадают, то эти процессы требуют большей активности и времени, чем в случае конгруэнтного слова, у которого артикуляция цвета шрифта совпадает с артикуляцией самого слова. Различия в трудности артикуляции цвета шрифта конгруэнтного и неконгруэнтного слова «красный» являются главной причиной, вызывающей появление временного эффекта Струпа на данное слово. В задаче 1 этот эффект локализован на латентности 370-444 мс, о чем свидетельствует физиологический эффект Струпа на это же слово, наблюдаемый в компоненте Р300 в отведении Т3.
Гипотеза о преимущественно артикуляторной природе временного эффекта Струпа позволяет объяснить его появление в задаче 2 на слово «зеленый». Это объяснение основано на сделанном выше предположении о том, что в задачу 2 включен процесс предварительной идентификации слов «зеленый» и «красный». Данное предположение позволило объяснить эффект фактора «подавление ответа», т. е. значительное увеличение ВР в ответ на предъявление слова «зеленый» в задаче 2. Однако оно позволяет также объяснить появление значимого временного эффекта Струпа в ответ на предъявление этого слова в данной задаче. Этот эффект может быть обусловлен тем, что в процессе предварительной идентификации стимула «зеленый» на ранней стадии его графическая форма неосознанно артикулируется, что дополнительно усложняет (для неконгруэнтного слова) или облегчает (для конгруэнтного слова) произвольную артикуляцию цвета шрифта этого стимула на поздней латентности. Это объясняет значимое увеличение временного эффекта Струпа в ответ на предъявление слова «зеленый»в задаче 2.
Отвечая на вопрос о том, на какой латентности локализован временной эффект Струпа в ответ на предъявление слова «зеленый» в задаче 2, следует также учесть, что физиологический эффект Струпа на слово «зеленый» возникает уже на латентности 49-63 мс. Он наблюдается в правой затылочной области мозга (в отведении О2) и связан, по-видимому, с процессом обнаружения конфликта между начавшимся восприятием цвета шрифта и начинающейся непроизвольной артикуляцией начертания стимула «зеленый».
Аналогичным образом можно объяснить физиологический эффект Струпа на латентности 56-61 мс в ответ на предъявление слова «красный» в правом височном отведении Т6 в задаче 2. В этой задаче не требовалось целенаправленно выяснять цвет шрифта данного слова. Тем не менее, процесс обнаружения конфликта между цветом шрифта слова «красный» и артикуляцией его начертания может осуществляться неосознанно и непроизвольно. Описанный процесс вносит определенный вклад во временной эффект Струпа, который в задаче 2 оказывается локализованным (т. е. связанным с процессами) не только на поздней (250 и более мс), но и на ранней (около 50 мс) латентности опознавательного действия. Это еще раз подтверждает известное утверждение о том, что психофизиологический механизм эффекта Струпа в значительной степени зависит от решаемой испытуемым задачи.
Таким образом, проведенное в данной работе совмещение экспериментальной парадигмы Струпа и «Стоп-сигнала», а также одновременная регистрация показателей ВР и ВП позволили получить ряд новых экспериментальных данных о возможных механизмах эффекта Струпа и эффекта фактора «подавление ответа». Было обнаружено, что временной эффект Струпа (т. е. влияние на ВР фактора «конгруэнтность стимулов») сопровождается физиологическим эффектом Струпа в определенных отведениях и компонентах ВП. Полученные результаты были достаточно полно объяснены с позиций сферической нейросетевой модели опознавательного действия (Беспалов, 2014; Соколов, Вайткявичус, 1989; Фомин, Соколов, Вайткявичус, 1979).
Выводы
Влияние на ВР и ВП фактора «подавление ответа», т. е. временной и физиологический эффекты данного фактора связаны с включением в задачу 2 (в ней подавляется ответ на слово «красный») процесса предварительной идентификации (оценки) тестовых стимулов как слов «зеленый» или «красный». С этими словами в задаче 2 с помощью инструкции соотнесены разные последующие процессы - выделение цвета шрифта у слова «зеленый» и отказ от ответа на слово «красный». Процесс предварительной идентификации стимулов в задаче 2 осуществляется на основе оценки их графической конфигурации (о чем свидетельствуют различия ВП в компоненте Р50) и последующей внутренней артикуляции графической «оболочки» (написания) стимульного слова (различия в компоненте N100). Предварительная идентификация стимулов в задаче 2 требует дополнительной активности и времени, что приводит к увеличению ВР (на 113 мс) в ответ на предъявление слова «зеленый», на которое в этой задаче давался двигательный ответ.
Влияние на ВРи ВПфактора «конгруэнтность стимулов» зависит от задачи. В задаче 1 оно связано с несовпадением (конфликтом) между процессом внутренней артикуляции воспринимаемого (с помощью нейронов-детекторов зрительной коры) цвета шрифта неконгруэнтного слова и процессом его непроизвольного чтения. Эти процессы протекают на латентности более 250 мс, о чем свидетельствует значимый физиологический эффект Струпа, наблюдаемый в левом височном отведении Т3. В задаче 2 эффект Струпа локализован как на ранней (около 50 мс), так и на поздней (около 450 мс) латентности опознавательного действия. На ранней латентности он обусловлен обнаружением конфликта между сенсорно воспринимаемым (с помощью нейронов-предетекторов в латеральных коленчатых телах) цветом шрифта тестового стимула и его предварительной идентификацией как слова «зеленый» или «красный» (путем выделения графической формы слова и, возможно, ее непроизвольной артикуляции). На поздней латентности в задаче 2 задержка ВР и уменьшение амплитуды ВП на неконгруэнтное слово «зеленый» возникает (как и в задаче 1) потому, что артикуляция цвета его шрифта не совпадает с непроизвольной артикуляцией графической формы слова.
Финансирование
Работа выполнена при поддержке гранта РФФИ № 13-06-00553а.