В недавно опубликованной книге «Облава на м-ра Альберта» (Driving Mr. Albert) рассказана подлинная история патологоа натома Томаса Харви, который в 1955 г. произвел вскрытие Альберта Эйнштейна. Выполнив работу, Харви самым непочтительным образом забрал мозг ученого домой, где в течение 40 лет хранил его в пластиковой банке с дезинфицирующей жидкостью. Время от времени патологоанатом отдавал маленькие срезы мозговой ткани исследователям из разных частей света, пытавшимся выяснить причины гениальности Эйнштейна. Когда Харви перевалило за 80, он погрузил остатки мозга в багажник своего «Бьюика» и повез возвращать их внучке гения.
Одной из тех, кто изучал срезы мозговой ткани Эйнштейна, была Мэриан Даймонд (Marian C. Diamond) — авторитетный гистолог из Калифорнийского университета в Беркли. Она установила, что числом и размерами нервных клеток (нейронов) головной мозг великого физика ничем не отличается от мозга обычного человека. Но в ассоциативной области коры, ответственной за высшие формы мыслительной деятельности, Даймонд обнаружила необычайно большое количество вспомогательных элементов нервной ткани — клеток нейроглии (глии). В мозге Эйнштейна их концентрация была намного больше, чем в голове среднестатистического Альберта.
Любопытное совпадение? Возможно. Но сегодня исследователи получают все больше данных, указывающих на то, что глиальные клетки играют гораздо более важную роль в деятельности мозга, чем предполагалось ранее. Долгие десятилетия все внимание физиологов было сосредоточено на нейронах — главных, по их мнению, приемопередатчиках мозга. Хотя глиальных клеток в 9 раз больше, чем нейронов, ученые отводили им скромную роль элементов, поддерживающих жизнедеятельность мозга (транспорт питательных веществ из кровенос-
ОБЗОР: ГЛИЯ
• Долгие десятилетия ученые полагали, что единственные клетки в головном мозге и других отделах нервной системы, способные к обмену сигналами, - это нейроны. Клеткам глии отводилась скромная роль вспомогательных элементов нервной ткани.
• Современные исследования показывают, что клетки глии обмениваются и с нейронами, и между собой посланиями о нейронной активности. Они способны изменять нейронные сигналы на уровне синаптических контактов между нейронами и влиять на образование синапсов.
• Таким образом, глия может играть решающую роль в процессах обучения и памяти, а также участвовать в восстановлении поврежденных нервов.
ных сосудов в нейроны, поддержание нормального баланса ионов в мозге, обезвреживание болезнетворных микробов, ускользнувших от преследования иммунной системы, и т.д.). Тем временем нейроны, поддерживаемые глией, общались друг с другом через крошечные контактные точки (синапсы) и формировали сложнейшие сети соединений, благодаря которым мы думаем, вспоминаем прошлое или испытываем радость.
Неизвестно, как долго просуществовала бы еще такая модель устройства мозга, если бы не недавно обнаруженные свидетельства того, что на протяжении всей жизни человека (от периода эмбрионального развития и до глубокой старости) нейроны и глия ведут весьма оживленный диалог. Глия влияет на образование синапсов и помогает мозгу определять, какие нервные связи усиливаются или ослабевают с течением времени (такие изменения напрямую связаны с процессами общения и долгосрочной памяти). Последние исследования показали, что глиальные клетки общаются идруг с другом, влияя на деятельность мозга в целом. Нейробиологи с большой осторожностью наделяют глию новыми полномочиями. Однако можно вообразить, какое волнение они испытывают при мысли о том, что большая часть нашего мозга почти не изучена и, следовательно, может еще раскрыть множество тайн.
ГЛИАЛЬНЫЕ КЛЕТКИ ОБЩАЮТСЯ С НЕЙРОНАМИ
Мы представляем себе нервную систему в виде переплетения проводов, соединяющих нейроны. Каждый нейрон снабжен одним длинным отростком — аксоном, переносящим электрические сигналы от тела нейрона к расширенным участкам на его конце — аксонным терминалям. Каждая терминаль высвобождает в синаптическую щель молекулы химического посредника — нейротрансмиттера, которые достигают соответствующих рецепторов на коротких ветвящихся отростках (дендритах) соседнего нейрона. Пространства между нейронами и аксонами заполнены массой разнообразных клеток глии. К тому времени, как скончался Эйнштейн, нейробиологи уже подозревали, что глиальные клетки принимают участие в переработке информации, но доказательств у них не было. В конце концов они оставили глию в покое.
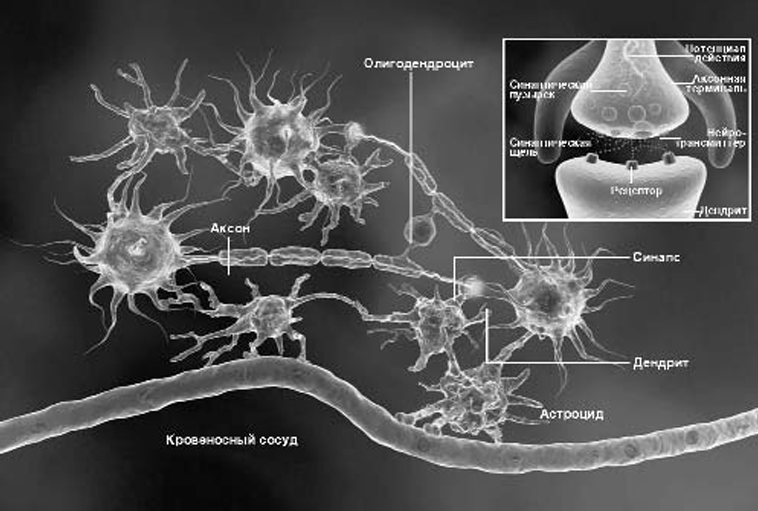
Глия и нейроны работают в головном и спинном мозге согласованно. Нейрон посылает по аксону сигнал, который через синаптическую щель достигает дендрита другой нервной клетки. Астроциты поставляют нейронам питательные вещест ва, а также окружают синапсы и регулируют их деятельность. Олигодендроциты вырабатывают миелин и образуют во круг аксонов изолирующие миелиновые оболочки. Когда электрический сигнал (потенциал действия) достигает аксон ной терминали (врезка), он заставляет пузырьки с нейротрансмиттером (сигнальными молекулами) перемещаться к мембране и высвобождать свое содержимое в синаптическую щель. Молекулы нейротрансмиттера диффундируют через узкую синаптическую щель к дендритным рецепторам. Точно так же осуществляется передача нервных сигналов и в периферической нервной системе, но здесь миелиновые оболочки вокруг аксонов образуют шванновские клетки.
Причина того, что исследователи не смогли обнаружить обмен сигналами между глиальными клетками, отчасти была связана с несовершенством методик. Но главными виновниками неудач были сами нейробиологи, ошибочно считавшие, что если клетки глии наделены способностью к общению, то обмениваться информацией они должны точно так же, как и нейроны, — с помощью электрических сигналов. Предполагалось, что клетки глии тоже должны генерировать электрические импульсы (потенциалы действия), стимулирующие выброс в синаптическую щель нейротрансмиттеров, которые, в свою очередь, вызывают импульсы в других клетках. Исследователи обнаружили, что глиальные клетки обладают несколькими типами ионных каналов, ответственных за генерирование электрических сигналов в аксонах, но они предположили, что эти каналы нужны глии просто для того, чтобы чувствовать уровень активности соседних нейронов. Было установлено, что мембрана глиальных клеток не обладает свойствами, необходимыми для проведения потенциалов действия. Нейробиологи, однако, упустили из виду одно обстоятельство, которое удалось обнаружить только благодаря современным методам исследования: глиальные клетки передают друг другу сообщения с помощью химических, а не электрических сигналов.
В середине 1990-х гг., ученые обнаружили в мембранах глиальных клеток рецепторы, реагирующие на разнообразные химические вещества, включая и нейротрансмиттеры. Это открытие навело их на мысль, что клетки глии способны общаться друг с другом с помощью сигналов, которые не распознаются нервными клетками.
Экспериментально было установлено, что показателем активации глиальных клеток служит поглощение ими кальция. На основании этого наблюдения исследователи разработали метод, позволяющий визуально определять, обладают ли терминальные шванновские клетки (один из типов глиальных клеток, окружающих синапсы в области контакта нервов с мышечными клетками) чувствительностью к нервным сигналам, приходящим к этим синапсам. Было установлено, что шванновские клетки действительно реагируют на синаптические импульсы, и что такая реакция сопровождается проникновением в них ионов кальция.
Но ограничивается ли участие глии в нервных процессах только «подслушиванием» нейронных переговоров? Ведь шванновские клетки окружают аксоны как в области синапсов, так и по ходу нервов в разных частях тела, а глиальные клетки другого типа (олигодендроциты) образуют оболочки вокруг аксонов в центральной нервной системе (т.е. в головном и спинном мозге). Сотрудники лаборатории Национального института здравоохранения США решили выяснить, способна ли глия отслеживать и нервные сигналы, распространяющиеся по аксонам в нервных цепях. И если такое общение между глией и нейронами существует, какие механизмы лежат в его основе и, что еще важнее, как влияют на работу глиальных клеток «подслушанные» ими нервные сообщения?
Чтобы ответить на эти вопросы, мы культивировали сенсорные нейроны (клетки дорсально-корешкового ганглия, ДКГ) мыши в специальных лабораторных чашках с электродами, с помощью которых можно было вызывать потенциалы действия в аксонах. В одни чашки с нейронами мы добавили шванновские клетки, в другие — олигодендроциты. Необходимо было одновременно контролировать активность и аксонов, и глии. За активностью нервных и глиальных клеток мы следили визуально, вводя в них краситель, который при связывании с ионами кальция должен был флуоресцировать. Когда по аксону пробегает нервный импульс, потенциалозависимые ионные каналы в нейронной мембране открываются, и ионы кальция проникают в клетку. Следовательно, распространение импульсов по аксонам должно сопровождаться зелеными вспышками внутри нейронов. По мере роста концентрации кальция в клетке флуоресценция должна становиться ярче. Ее интенсивность можно измерить с помощью фотоэлектронного умножителя, а искусственно окрашенные изображения светящейся клетки воспроизвести в реальном времени на экране монитора. Если глиальные клетки реагируют на нервные сигналы и поглощают в это время ионы кальция из окружающей среды, они тоже должны засветиться — только немного позднее, чем нейроны.
Сидя в затененной комнате и напряженно вглядываясь в экран монитора, мы с биологом Бетом Стивенсом (Beth Stevens) собирались приступить к эксперименту, на подготовку которого у нас ушло несколько месяцев. На включение стимулятора нейроны ДКГ тут же отреагировали изменением цвета: по мере увеличения концентрации кальция в их аксонах они превратились из синих в зеленые, затем — в красные и, наконец, побелели. Сначала ни в шванновских клетках, ни в олигодендроцитах никаких изменений не обнаружилось, но спустя 15 долгих секунд они, подобно елочным лампочкам, начали загораться. Клетки глии почувствовали, что по аксонам пробегают импульсы, и отреагировали на это событие увеличением концентрации кальция в цитоплазме.
ГЛИАЛЬНЫЕ КЛЕТКИ ОБЩАЮТСЯ ДРУГ С ДРУГОМ
Нам удалось показать, что глия способна распознавать импульсную активность в аксонах, реагируя на нее поглощением кальция. В нейронах он активирует ферменты, ответственные за выработку нейротрансмиттеров. Вполне вероятно, что поступление кальция в глиальные клетки также вызывает активацию ферментов, связанных с развитием какой-то реакции. Но какой?
Исследование еще одного типа глиальных клеток — астроцитов, транспортирующих питательные вещества из капилляров в нервные клетки и поддерживающих оптимальный уровень ионов, необходимый для генерирования нервных импульсов в окружающей нейроны среде (включая и удаление избытка нейротрансмиттеров и ионов, высвобождаемых нейронами во время импульсации), поможет ответить на этот вопрос. В 1990 г.
Стивен Смит из Йельского университета показал, что если в культуру астроцитов добавить нейротрансмиттер глутамат, концентрация кальция в клетках резко возрастает. Клетки ведут себя так, словно только что произошел выброс нейротрансмиттера из нейрона и они горячо обсуждают друг с другом вызвавшую его импульсацию нейронов.
Некоторые нейробиологи пытались выяснить, не явилось ли общение глиальных клеток следствием простого перемещения ионов кальция или связанных с ним сигнальных молекул из одного астроцита в соседний через соединяющие их открытые ворота. В 1996 г. исследователи из Университета штата Юта опровергли это предположение. С помощью острого микроэлектрода они разрезали слой астроцитов в культуре на две части, оставив между ними зазор, не содержавший клеток и разделявший популяцию астроцитов. Когда концентрация кальция в клетках по одну сторону разреза возрастала, то же происходило и по другую сторону. Таким образом выяснилось, что астроциты посылали друг другу сигналы через внеклеточную среду.
АТФ КАК ХИМИЧЕСКИЙ ПОСРЕДНИК
Выявленные закономерности привели исследователей в замешательство. Коммуникация глиальных клеток, как и нейронов, контролируется токами кальция. Однако если изменения его уровня в нейронах вызывают электрические импульсы, то в глии — нет. Возникает вопрос: не было ли инициировано перемещение ионов кальция в глию каким-то другим электрическим феноменом? А если нет, то какова природа механизма?
Когда нейробиологи экспериментировали с глией, в их поле зрения постоянно попадала знакомая всем молекула аденозинтрифосфата (АТФ). Будучи основным источником энергии в живых клетках, АТФ обладает многими признаками, благодаря которым прекрасно подходит на роль химического посредника между клетками. В окружающей среде он содержится в больших количествах, а во внеклеточном пространстве его мало. Благодаря небольшим размерам молекула способна к быстрой диффузии и легко разрушается ферментами. Более того, АТФ присутствует в аксонных терминалях, где обычно и хранятся молекулы нейротрансмиттеров, и может высвобождаться в синаптическую щель.
В 1999 г. исследователи из Университета штата Юта показали, что при возбуждении астроциты выбрасывают в окружающую среду АТФ. Затем он связывается рецепторами на соседних астроцитах, заставляя открываться ионные каналы и способствуя перемещению кальция внутрь клеток. В свою очередь, повышение уровня кальция в клетках заставляет их высвобождать во внеклеточную среду новые порции АТФ — так в популяции астроцитов инициируется цепная реакция, связанная с изменением внутриклеточного уровня кальция и опосредованная АТФ.
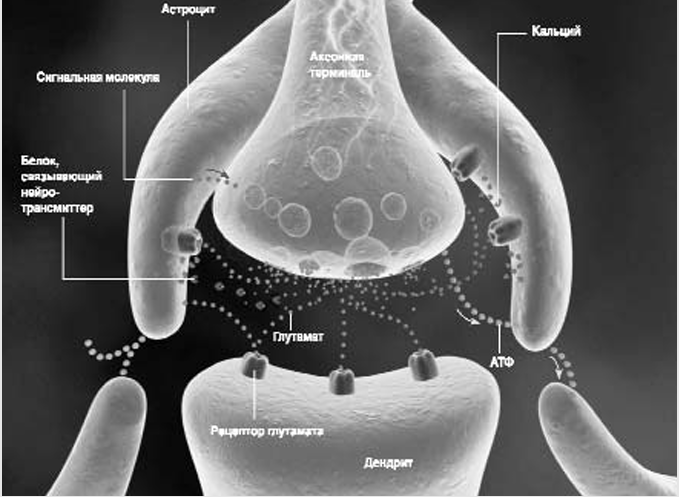
Астроциты регулируют синаптическую передачу сигнала несколькими способами. Аксон передает нервный сигнал дендриту за счет выброса нейротрансмиттера (зеленый) - в данном случае глутамата. Кроме того, аксон высвобождает АТФ (желтый). Эти соединения вызывают перемещение кальция (фиолетовый) внутрь астроцитов, что побуждает их вступить в общение друг с другом за счет высвобождения собственного АТФ. Астроциты могут усилить передачу нервного сигнала с помощью выброса такого же нейротрансмиттера (глутамата) или ослабить сигнал путем поглощения нейротрансмиттера или выброса связывающих его белков (синие). Кроме того, астроциты могут выделить сигнальные молекулы (красные), которые заставят аксон увеличить или уменьшить выброс нейротрансмиттера, когда он возобновит импульсацию. Модификация связей между нейронами - один из способов, с помощью которых головной мозг корректирует свои реакции на раздражители по мере накопления опыта, - так происходит процесс обучения. В периферической нервной системе синапсы окружены не астроцитами, а шванновскими клетками.
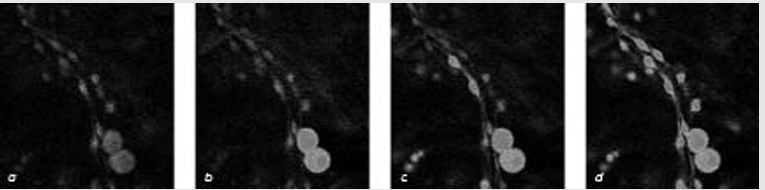
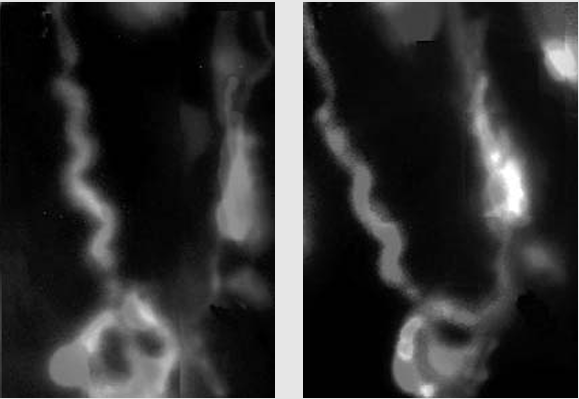
Кадры фильма (окраска искусственная), отснятого с помощью лазерной конфокальной микроскопии, показывают, что глиальные клетки способны реагировать на обмен сигналами между нейронами. Сенсорные нейроны (две крупные клетки диаметром 20 микрон) (a) и более мелкие шванновские клетки были помещены в культуральную среду, содержащую ионы кальция. В клетки вводился краситель, который при связывании ионов кальция начинал флуоресцировать. Стимуляция нейронов электрическим током небольшого напряжения заставляла их генерировать распространяющиеся по аксонам (длинные линии) импульсы (потенциалы действия). Нейроны при этом тут же начинали флуоресцировать (b), что указывает на проникновение в них кальция через открывшиеся ионные каналы в мембране. Спустя 12 секунд (с), когда нейроны продолжали генерировать разряды, начали светиться и шванновские клетки - т.е. в ответ на распространение нервных сигналов по аксонам они тоже начали поглощать кальций из внеклеточной среды. Через 18 секунд после этого (d) свечение охватило еще большее количество глиальных клеток, почувствовавших сигналы нейронов. Как видно из представленной серии кадров, глия «подслушивала» нейронные сообщения по всему ходу комму. никационных линий, а не только в области синапсов, где присутствовал нейротрансмиттер.
В результате наблюдений родилась модель, объясняющая способность околоаксонной глии распознавать нейронную активность и затем передавать сообщения другим глиальным клеткам, окружающим синапс. Импульсация нейронов побуждает глиальные клетки, окружающие аксон, высвобождать АТФ, который вызывает поглощение кальция соседними глиальными клетками. Это стимулирует выброс новых порций АТФ, что активирует передачу сообщения по длинной цепочке глиальных клеток иногда на значительное расстояние от нейрона, инициировавшего всю последовательность данных событий. Но каким образом глиальным клеткам, участвовавшим в нашем эксперименте, удавалось распознавать импульсацию нейронов — ведь аксоны не образуют с глией синаптических контактов и в области синапсов не было никаких глиальных клеток? Участием нейротрансмиттеров такое явление объяснить нельзя: из аксонов они не диффундируют. Быть может, его причиной был АТФ, способный каким-то образом просачиваться из аксонов?
Для проверки гипотезы мы решили провести электрическую стимуляцию чистых культур аксонов ДКГ и последующий химический анализ культуральной среды. Воспользовавшись ферментом, ответственным за свечение брюшка у жуков-светляков (эта реакция требует участия АТФ), мы наблюдали свечение среды во время распространения импульса по аксонам, что свидетельствовало о высвобождении из них АТФ. Затем мы добавили в культуру шванновские клетки, также начинавшие светиться после того как по аксонам пробегали потенциалы действия. Но когда мы добавили в среду фермент апиразу, быстро разрушающую АТФ и не дающую ему достигнуть шванновских клеток, глия во время импульса- ции аксонов оставалась темной. Таким образом, содержание кальция в шванновских клетках не менялось, т.к. они не получали АТФ-сигнала.
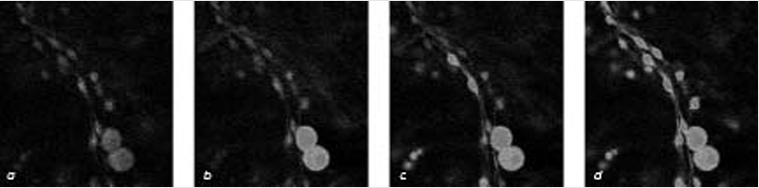
Как общаются глиальные клетки? В культуральную среду, содержавшую кальций, помеща лись астроциты (а) и сенсорные нейроны. После того как под влиянием электрической стиму ляции нейроны принялись генерировать распространяющиеся по аксонам (зигзаги молний) (b) импульсы (потенциалы действия), глия начала флуоресцировать – признак того, что гли альные клетки отреагировали на это событие поглощением кальция. Спустя 10 и 12,5 секунд (с и d) по всей популяции астроцитов прокатились две огромные волны проникновения каль ция внутрь клеток. О росте концентрации кальция в астроцитах свидетельствует изменение их цвета: вначале они были зелеными, затем стали синими и наконец красными.
АТФ, высвободившийся из аксонов, стимулировал транспорт кальция внутрь шванновских клеток. С помощью биохимического анализа и цифровой микроскопии нам удалось показать, что в результате этого события сигнальные молекулы перемещаются от клеточной мембраны к ядру и включают здесь различные гены. Таким образом, мы обнаружили поразительный факт: генерируя импульсы, призванные обеспечить общение с другими нейронами, нервная клетка и ее аксон влияют на считывание генов в глиальной клетке и изменяют тем самым ее поведение.
АКСОНЫ ОПРЕДЕЛЯЮТ СУДЬБУ ГЛИАЛЬНЫХ КЛЕТОК
Какие функции глии могут контролировать гены, включенные АТФ? Приказывают ли они глиальным клеткам оказывать влияние на окружающие их нейроны? Исследователи попытались ответить на вопрос, обратив внимание на процесс, способствующий образованию миелиновой изолирующей оболочки вокруг аксонов. Благодаря ей аксоны способны проводить нервные импульсы с огромной скоростью на значительные расстояния. Ее образование позволяет малышу все крепче удерживать голову в вертикальном положении, а разрушение вследствие некоторых болезней (например, рассеянного склероза) превращает человека в инвалида.
Мы решили выяснить, как незрелая шванновская клетка, расположенная на аксоне в периферической нервной системе плода или младенца, узнает, нуждается ли отросток в миелинизации и когда нужно приступить к его «пеленанию» миелином. Или, напротив, следует ли ей превратиться в клетку, которая не будет сооружать миелиновую оболочку? Могут ли аксональные нервные импульсы или высвобождение АТФ влиять на выбор шванновской клетки? Мы обнаружили, что шванновские клетки в культуре пролиферировали медленнее в том случае, когда окружали импульсирующие, а не молчащие аксоны. Более того, они приостанавливали свое развитие и прекращали выработку миелина. Добавление АТФ вызывало такое же действие.
ГЛИЯ КОНТРОЛИРУЕТ СИНАПСЫ
Бен Баррес из Стэнфордского университета обнаружил, что если выращивать нейроны из сетчатки крысы в лабораторной культуре, не содержавшей астроцитов, синапсов на нейронах образуется очень мало. Когда же он добавил в культуру астроциты или просто среду, в которой прежде находились астроциты, синапсы появились в большом количестве. Затем он обнаружил присутствие в среде двух химических веществ, высвобождаемых астроцитами для стимуляции образования синапсов, - жировой комплекс под названием ароЕ/холестерин и белок тромбоспондин. Немного позднее Ле Тиан и Уэсли Томпсон из Техасского университета в Остине изучали мышей, которым вводились вещества, заставлявшие флуоресцировать шванновские клетки, что позволило им наблюдать за деятельностью глиальных клеток в области контактов нервов с мышечными волокнами. После того как исследователи перерезали подходящий к мышце аксон, нервно-мышечный синапс исчезал, но на его «мышечной стороне» оставалась группа рецепторов нейротрансмиттера. Исследователям было известно, что аксон вновь сможет прорасти к покинутым им рецепторам. Но как он найдет к ним путь? Следя за флуоресценцией, Томпсон увидел, что шванновские клетки, окружавшие интактные синапсы, почувствовали, что синапс-сосед оказался в беде. Тогда они дружно выпустили в его сторону отростки, дотянулись ими до поврежденного синапса и образовали своего рода мостик, по которому аксон мог послать к своему синапсу новую проекцию (см. фото). Эти данные указывают на то, что глия помогает нейронам определить место, где нужно образовывать синаптические соединения. Сегодня нейробиологи пытаются использовать такую способность глии для лечения повреждений спинного мозга: они трансплантируют шванновские клетки в поврежденные участки спинного мозга лабораторных животных.
Глия способна управлять образованием синапсов. Нейробиологи перерезали у мыши нерв, подходивший к нервномышечному синапсу. Спустя два дня (левый снимок) отростки шванновских клеток (темнокрасный цвет) образовали в области разреза (отмечен стрелкой) своего рода мостик. А еще через пару дней (правый снимок) аксон (зеленый) направил по этому мостику новую проекцию к синапсу.

В мозге Эйнштейна концентрация клеток нейроглии была намного больше, чем в голове среднестатистического Альберта
Витторио Галло (Vittorio Gallo) из соседней лаборатории НИЗ, изучая олигодендроциты, образующие миелиновые оболочки вокруг аксонов в головном мозге, обнаружил совершенно иную картину. АТФ не угнетал пролиферацию клеток, но аденозин (вещество, в которое превращается молекула АТФ после отщепления от нее остатков фосфорной кислоты) стимулировал созревание клеток и выработку миелина.
Понимание механизмов миелинизации имеет важнейшее значение. Болезни, сопровождающиеся разрушением миелиновой оболочки, каждый год уносят тысячи человеческих жизней и вызывают паралич и слепоту. Неизвестно, какой фактор инициирует миелинизацию, но аденозин стал первым веществом «аксонального происхождения», у которого была выявлена способность стимулировать этот процесс. Аденозин высвобождается из аксонов в ответ на распространение импульсов, и это означает, что электрическая активность мозга действительно влияет на процесс миелинизации. Подобные открытия помогут исследователям вести поиск средств для лечения болезней демиелинизации. Возможно, эффективными окажутся лекарства, напоминающие своей химической структурой аденозин. И не исключено, что добавление аденозина в культуру стволовых клеток превратит их в миелинизирующие глиальные клетки, которые можно будет использовать в качестве трансплантатов.
ВЫРЫВАЯСЬ ИЗ ПУТ НЕЙРОННЫХ СЕТЕЙ
Ограничивается ли участие глии в регуляции нейронных функций образованием вокруг аксонов миелиновой оболочки? По-видимому, нет. Ришар Робитайль (Richard Robitaille) из Монреальского университета обнаружил, что величина электрического потенциала, возникающего в мышце лягушки под влиянием стимуляции синапса, увеличивалась или уменьшалась в зависимости от того, какие химические вещества он вводил в шванновские клетки, окружающие этот синапс. Когда исследователь прикасался к сетчатке крысы, посылаемые глией «кальциевые сигналы» изменяли частоту импульсации зрительных нейронов. А Майкен Недергард (Maiken Nedergaard) из Нью-Йоркского медицинского колледжа, изучавший срезы гиппокампа крысы, наблюдал усиление электрической активности синапсов в то время, когда окружающие астроциты увеличивали поглощение кальция. Подобные изменения эффективности синапсов специалисты рассматривают в качестве главного фактора пластичности нервной системы, т.е. ее способности изменять реакции на основании прошлого опыта, и глия, таким образом, может играть важную роль в клеточных процессах обучения и памяти.
В связи с описанными выше наблюдениями остро встает одна проблема. Поглощение кальция распространяется по всей популяции астроцитов, подобно прокатывающимся по стадиону волнам взявшихся за руки болельщиков. Такая дружная реакция необходима для управления работой всей группы клеток, но она слишком груба для передачи сложных сообщений. Принцип «все как один!» может оказаться полезным для координации общей активности мозга во время цикла сон-бодрствование, но чтобы входить во все тонкости переработки информации, глиальные клетки должны уметь «переговариваться» и со своими непосредственными соседями.
Исследователи предполагают, что нейроны и клетки глии способны вести друг с другом беседы и более «интимного» свойства. Экспериментальные методы, которыми располагали в то время нейробиологи, не позволяли им апплицировать нейротрансмиттеры в таких ничтожно малых дозах, которые могли бы воспроизвести истинные «переживания» астроцита, находящегося рядом с синапсом. Филипу Хейдону (Philip G. Haydon) из Пенсильванского университета удалось добиться этого только в 2003 г. с помощью современного лазерного метода аппликации нейротрансмиттеров. Он стимулировал в срезах гиппокампа выброс такого ничтожного количества глутамата, которое мог обнаружить только один-единственный астроцит. Хейдон наблюдал при этом, что астроцит посылает специфические кальциевые сигналы лишь небольшому числу окружающих его астроцитов. Исследователь предположил, что наряду с «кальциевыми волнами», оказывающими широкомасштабное воздействие, «между астроцитами существуют и близкодействующие связи». Иными словами, разрозненные цепочки астроцитов в головном мозге координируют свою активность в соответствии с активностью нейронных цепей.
Описанные выше открытия позволили Хейдону сформулировать рабочую гипотезу, согласно которой обмен сигналами помогает астроцитам активировать нейроны, чьи аксоны оканчиваются на сравнительно большом от них расстоянии, а также утверждать, что такая активация способствует высвобождению нейротрансмиттеров из отдаленных синапсов. Это позволяет астроцитам регулировать готовность отдаленных синапсов к изменению своей силы, что является клеточной основой процессов памяти и обучения.
Результаты исследований, представленные на ежегодном съезде Общества нейробиологии в ноябре 2003 г., подкрепляют данную гипотезу и даже свидетельствуют об участии глии в образовании новых синапсов (врезка на с. 85). Исследователи из Стэнфордского университета, установили, что выращенные в культуре нейроны крысы в присутствии астроцитов образуют большее количество синапсов. Впоследствии они обнаружили, что белок тромбоспондин, предположительно астроцитарного происхождения, выполняет функцию химического посредника и стимулирует образование синапсов. Чем большее количество этого белка ученые добавляли в культуру астроцитов, тем больше синапсов появлялось на нейронах. Возможно, тромбоспондин отвечает за связывание белков и других соединений, необходимых для образования синапсов во время роста молодых нервных сетей и, следовательно, может участвовать в модификации синапсов, когда эти сети подвергаются старению.
Будущие исследования расширят наши представления о влиянии глии на нейронную часть головного мозга. Возможно, нейробиологам удастся доказать, что наша память (или ее клеточный аналог — такой, как долговременная потенциация) зависит от функционирования синаптических астроцитов. Не исключено также, что будет установлено, каким образом сигналы, передаваемые по цепочкам астроцитов, оказывают влияние на отдаленные синапсы.
Сравнение головного мозга показывает, что чем более высокое положение занимают животные на «эволюционной лестнице», тем выше у них соотношение между числом глиальных клеток и нейронов. Хейдон предполагает, что увеличение связности астроцитов может повышать способность животных к обучению. Не исключено, что высокие концентрации глиальных клеток в мозге, а возможно, и наличие в нем более «действенной» глии, и превращают некоторых людей в гениев. Эйнштейн учил нас думать нетрадиционно. Его примеру последовали ученые, дерзнувшие «выпутаться» из нейронных сетей и решившие наконец выяснить, какое участие в переработке информации принимает нейроглия.