Введение
Хотя хорошо известно, что цветовое зрение человека основано на сравнении количества фотонов, поглощаемых тремя типами колбочек (Dartnall et al., 1983; Schnapf et al., 1987; Измайлов и др., 1989), однако процессы, лежащие в основе цветовосприятия на пострецепторных стадиях, до сих пор остаются недостаточно изученными. Одна из основных проблем – несоответствие красно-зеленой и сине-желтой субъективных, или феноменологических, осей цветового пространства человека цветовым оппонентным каналам, существование которых было показано на первичных стадиях обработки сигналов в зрительной системе приматов. Феноменологическая красно-зеленая ось разделяет цвета, имеющие желтые и синие оттенки, и проходит через точки в цветовом пространстве, соответствующие чистому красному и чистому зеленому цветам, а сине-желтая ось разделяет цвета, имеющие красные и зеленые оттенки, и проходит через точки чистого желтого и чистого синего цветов.
Регистрация ответов нейронов головного мозга макак показала, что на первичных стадиях различение цветов зависит от двух оппонентных сигналов, образующихся на уровне сетчатки и наружного коленчатого тела (Derrington et al., 1984). Один из этих хроматических сигналов представляет собой разницу или скорее отношение сигналов, поступающих от длинноволновых колбочек (Long-wave sensitive cones, или L-колбочек) и от средневолновых колбочек (Middle-wave sensitive cones, или M-колбочек); этот сигнал поступает на карликовые ганглиозные клетки сетчатки и далее в парвоцеллюлярные слои НКТ. Второй хроматический сигнал представляет отношение сигналов, поступающих от коротковолновых колбочек (Short-wave sensitive cones, или S-колбочек) и комбинации сигналов от L-и М-колбочек; этот сигнал передается через малые бистратифицированные ганглиозные клетки сетчатки и далее кониоцеллюлярными слоями НКТ (Dacey, Lee, 1994). Этих два нейронных канала соответствуют горизонтальной и вертикальной осям диаграммы цветности МакЛаода-Бойнтона (рис. 1, левая панель), в настоящее время широко используемой для представления диапазона цветов, которые различает зрительная система человека (MacLeod, Boynton, 1979).
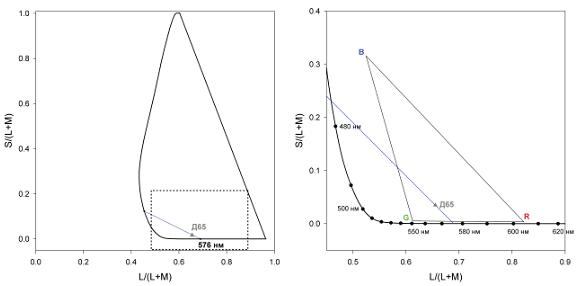
Рис. 1. Левая панель. Диаграмма цветностей МакЛаода-Бойнтона. Абсцисса представляет собой возбуждение L-колбочек по отношению к суммарному возбуждению L- и M-колбочек; ордината представляет возбуждение S-колбочек по отношению к суммарному возбуждению L- и M-колбочек. Сплошная линия показывает положение спектральных цветов и фиолетовых оттенков (верхняя часть диаграммы): все эти цвета представляют гамму цветов, различаемых наблюдателями с нормальным цветовым зрением. Д65 указывает положение стандартного МКО источника освещения Д65. Линия, проходящая через Д65 и 576 нм, является нашей предварительной оценкой сине-желтой линии, по литературным данным. Правая панель. Представлена область диаграммы, заключенная в прямоугольник из штриховой линии на левой панели. У диаграммы на этой панели изменен масштаб так, что линия Д65–576 нм проходит под углом -45 градусов к абсциссе. R, G и B показывают положения цветностей люминофоров монитора SONY: они определяют гамму цветностей, которую можно воспроизвести с помощью такого монитора. Линия, проходящая через точки Д65 и 576 нм, – наша предварительная оценка сине-желтой линии.
Хотя цветность – величина непрерывная в обоих направлениях диаграммы цветности (сигналы от трех колбочек дают независимые отношения, которые представлены двумя осями), мы группируем цвета, разделяя двухмерную диаграмму на ограниченный дискретный набор категорий, и таким образом наш феноменологический опыт сводит все цвета к ограниченному набору цветовых оттенков (Корж и др., 1991; Соколов, Вартанов, 1987). Однако сама фундаментальная проблема заключается в том, что нейронные каналы, существование которых показано в сетчатке и НКТ, не соответствуют феноменологическим сине-желтой и красно-зеленой осям пространства восприятия цветов. Сине-желтая ось – это линия, соединяющая чистый синий и чистый желтый цвета; она делит диаграмму цветности на области, содержащие цвета, обладающие красноватыми (выше оси) или зеленоватыми (ниже оси) оттенками. В пространстве МакЛаода-Бойнтона эта линия является наклонной и не соответствует ни одной из физиологических осей, или нейронных каналов, которые представлены на диаграмме цветности абсциссой и ординатой (рис. 1, правая панель). Нейронный механизм, лежащий в основе воспринимаемых цветовых категорий, до сих пор неизвестен и является одной из основных проблем современной когнитивной нейронауки. Немногочисленные эксперименты, предполагающие существование нейрофизиологических механизмов в задней нижней височной коре (Stoughton, Conway, 2008), не выдерживают критических замечаний (Mollon, 2009).
Более точное различение цветов на границе между категориями
Пороги цветоразличения не являются одинаковыми в разных участках диаграммы цветности МакЛаода-Бойнтона и других общепринятых диаграмм цветности, включая стандартную диаграмму МКО1931. Также не существует способа изменения масштаба осей, который бы сделал диаграмму равномерной с точки зрения равенства порогов цветоразличения. Существует два традиционных подхода к вопросу о том, где должно наблюдаться оптимальное цветоразличение. Один – традиционный – подход основан на наличии воспринимаемых цветовых категорий, а другой – на нейрофизиологии первых этапов обработки информации о цвете в зрительной системе.
В первом случае традиционно предполагается, что цветоразличение будет оптимально на границе между двумя цветовыми категориями. Можно провести классическую аналогию со слуховым восприятием: в английском языке в случае звонких согласных, таких, как b, d и g, слушатели не воспринимают малые изменения внутри каждой категории, но хорошо различают звуки на границе категорий (Liberman et al., 1957). В случае цветовых категорий в подавляющем большинстве исследований положительный результат был получен при изучении скорости различения или при применении надпороговых стимулов, а не при измерении порогов цветоразличения. При использовании набора стимулов в синей и голубой областях было показано, что русскоговорящие наблюдатели находят быстрее стимул, отличающийся от двух других одинаковых, если стимулы выбраны из разных категорий «голубой» и «синий» (Winawer et al., 2007). Англоговорящие наблюдатели, у которых для всех тех же цветов используется только один термин «blue», не показали более быстрого различения цветов, находящихся по разные стороны от границы «светлый blue» и «темный blue». В области переходных – от зеленых к синим – цветов также было зарегистрировано более короткое время реакции для стимулов, которые предварительно были уравнены по воспринимаемой яркости (Witzel et al., 2009). При измерении порогов цветоразличения, а не времени реакции (Roberson et al., 2009) не было получено улучшения цветоразличения, т. е. минимальных порогов, на границе категорий «синий» и «зеленый» для англоговорящих наблюдателей, а также не было получено разницы между английскими и корейскими наблюдателями при различении цветов на границе категорий, которые существуют в корейском языке и не существуют в английском языке.
В случае второго – нейрофизиологического – подхода наилучшее цветоразличение, или оптимальные пороги, должно наблюдаться в случае, если два стимула находятся на наиболее крутой части функции ответов нейронов, которые отвечают в процессе обнаружения. В этом случае минимальная разница между контрастом двух стимулов приводит к максимальной разнице в ответах таких нейронов. В случае яркостного контраста известно, что наиболее крутая часть амплитудной характеристики (зависимость величины реакции нейрона от логарифма относительной величины изменения интенсивности света) сдвигается в зависимости от адаптации (Бызов, Кузнецова, 1969). Максимальная контрастная чувствительность человека получена для стимулов, контраст которых является либо инкрементом, либо декрементом от текущего уровня фона, к которому адаптирована зрительная система, и снижается при удалении от уровня фона. Психофизические данные свидетельствуют также о том, что аналогичные процессы имеют место и в случае цветовых нейронных каналов: пороги цветоразличения минимальны для цветов, близких к цвету адаптирующего фона (Krauskopf, Gegenfurtner, 1992; Loomis, Berger, 1979; Miyahara et al., 1993; Раутиан, Соловьева, 1954). Рассмотрим парвоцеллюлярный канал, который получает входы противоположного знака от L- и M-колбочек. В этом случае нейрофизиологический подход говорит о том, что такой канал будет наиболее чувствителен, если сигналы от L- и M-колбочек сбалансированы, т. е. отношение возбуждений L- и M-колбочек при предъявлении стимулов не сильно отличается от отношения возбуждений L- и M-колбочек, возникающих при предъявлении адаптирующего фона. Когда отношение L:M отклоняется в большей степени от отношения, даваемого адаптирующим фоном, т. е. когда канал становится более поляризованным, его чувствительность уменьшается. Другими словами, функция ответов такого канала нелинейная. Де Валуа и другие (De Valois et al., 1967) регистрировали ответы нейронов НКТ макак и определяли возможности различения цветов отдельными нейронами. Они показали, что ответы цвето-оппонентных клеток имеют область оптимальной чувствительности, которая находится в области длины волны фона, к которому в данный момент адаптирована исследуемая клетка.
Различение в направлении, перпендикулярном к сине-желтой оси
Наши предыдущие эксперименты были направлены на исследование возможности наблюдателей сравнивать стимулы, предъявляемые одновременно на короткое время в разных участках поля зрения и на разном расстоянии друг от друга в парафовеа (Danilova, Mollon, 2010). В этих экспериментах был обнаружен следующий факт: различение цветов, которые находятся на линии, перпендикулярной сине-желтой оси, существенно улучшается в области границы между зеленоватыми и красноватыми цветами, т. е. различение цветов, расположенных по разные стороны от этой оси, происходит с большей точностью. Эта граница не соответствует условию равновесия ни L/M, ни S/(L+M) каналов, т. е. когда оппонентные входы этих каналов сбалансированы. Отсутствие такого результата в литературе может быть связано с тем, что мы получили этот эффект для цветов, близких к цветам, встречающимся в реальных ситуациях, а не для цветов, близких к монохроматическим.
Приведенные в настоящей статье результаты экспериментов в фовеа показывают, что наличие минимальных порогов цветоразличения в определенной области диаграммы цветности не является свойством парафовеального зрения: аналогичный эффект наблюдается также и в фовеа. Вместо двух стимулов, предъявляемых в парафовеа на расстоянии 3,7 угловых градуса между их центрами, в фовеа мы использовали классический стимул, применяемый при исследовании цветоразличения, – диск диаметром 2 угловых градуса, разделенный вертикально на две равные части (Wyszecki, Stiles, 1982). Для спецификации наших стимулов мы изменили масштаб диаграммы цветностей МакЛаода-Бойнтона таким образом, чтобы сине-желтая линия проходила под углом -45 градусов к абсциссе. Затем были выбраны пять точек на этой линии, через которые были проведены ортогональные линии под углом +45 градусов к абсциссе. На этих линиях были выбраны семь точек с обеих сторон сине-желтой линии, для которых измеряли пороги цветоразличения. В отдельных экспериментах также проводили феноменологические измерения вдоль тех же пяти линий. В этих сериях просили наблюдателей выбрать такой цвет, в котором нет оттенков красного или зеленого. Целью этих экспериментов было определение субъективной границы между категориями красноватых и зеленоватых цветов.
Во второй серии эксперимента мы проверяли, является ли обнаруженный нами эффект действительно сенсорным явлением или такой результат был получен, потому что для наблюдателей установить соответствие между категорией и ответом проще в районе точки равновесия, т. е. когда цвета находятся по разные стороны сине-желтой оси, так как в этом случае наблюдатель делает меньше ошибок. Это проверялось также при использовании процедуры вынужденного выбора, но варианта метода «одинаковый–разный» (Mollon, Cavonius, 1987). Преимущество этого метода в том, что наблюдатель не должен выносить суждение о цвете, а только решать, в каком из двух временных интервалов две половины диска были разные по цвету.
Эксперимент 1 (а, б). Методика
Наблюдатели. Все наблюдатели обладали нормальным цветовым зрением, проверка производилась с использованием компьютеризированного цветового теста Cambridge Colour Test (Regan et al., 1994). MВД и ДДМ (авторы статьи) являются опытными наблюдателями; родной язык МВД – русский, ДДМ – английский. Шесть наивных наблюдателей участвовали в эксперименте 1 а. У пяти из них родной язык – английский, у одного наблюдателя родным является немецкий язык.
Аппаратура и стимулы. Стимулы предъявляли на экране калиброванного электронно-лучевого монитора при использовании графической платы, которая имеет разрешение 15 бит на каждую электронную пушку. Наблюдатель ДДМ и все остальные наблюдатели, принимавшие участие в эксперименте 1а и 2, участвовали в экспериментах в Кембридже, где использовался монитор Mitsubishi Diamond Pro 2070 (частота смены кадра 92 Гц, разрешение 1024 х 768 пикселей) и графическая карта VSG 2/3 (Cambridge Research Systems, Rochester, UK). Наблюдатель МВД проходил тестирование в Санкт-Петербурге, где в установке использовали монитор SONY GDM-F500 (частота смены кадра 80 Гц, разрешение 1280 х 960 пикселей) и графическую карту VSG 2/5 (Cambridge Research Systems, Rochester, UK). Процедуры калибровки и алгоритмы для воспроизведения заданных цветов на экране электронно-лучевого монитора были стандартными и одинаковыми для двух экспериментальных установок (Cole, Hine, 1992; Smith, Pokorny, 1995). Для измерения спектров испускания люминофоров использовали спектрофотометр JETI SpecBos (JETI Technische Instrumente GmbH), для коррекции гамма-функции монитора использовали фотометр OptiCal (Cambridge Research Systems, Rochester, UK).
Для спецификации цвета стимулов мы использовали диаграмму цветностей МакЛаода-Бойнтона (рис. 1, левая панель), которая становится стандартной диаграммой среди ученых, занимающихся исследованием цветовосприятия и колориметрией (MacLeod, Boynton, 1979). Плоскость диаграммы цветности представляет плоскость равной яркости для стандартного наблюдателя, предложенного Джаддом в 1951 году. Яркость равна сумме сигналов L- и M-колбочек, L+M (Smith, Pokorny, 1975). Для построения диаграммы мы использовали кривые спектральной чувствительности, приведенные в статье ДеМарко и других авторов (DeMarco et al., 1992). Абсцисса диаграммы представляет ту часть яркости, которая определяется вкладом L-колбочек, т. е. L/(L+M), а ордината представляет возбуждение S-колбочек по отношению к комбинации возбуждений L- и M-колбочек, S/(L+M). В наших экспериментах мы использовали для фона значение L+M, соответствующее 10 кд/м-2 яркости, измеренной в единицах МКО1931. Яркость двух половин диска, служащих стимулами, была на 30% выше яркости фона в единицах L+M нашей диаграммы. Чтобы быть уверенными, что наблюдатели не используют субъективную яркость стимулов как один из признаков для различения цветов, в каждом предъявлении варьировали яркость каждой половины диска в пределах ± 5% от среднего значения в единицах L+M.
Поскольку масштаб оси S/(L+M) диаграммы МакЛаода-Бойнтона произвольный, мы выбрали его таковым, что линия, соединяющая чистый синий и чистый желтый цвета, имеет наклон -45 градусов (рис. 1, правая панель). Из литературных данных известно, что чистый синий и чистый желтый находятся близко к 476 нм и 576 нм соответственно (Burns et al., 1984; Jordan, Mollon, 1997; Nerger et al., 1995; Purdy, 1931; Webster et al., 2002). Наша линия проходит через точки 576 нм и 478 нм (линия Д65–576 нм). Мы выбрали именно такую линию, поскольку она проходит также и через точку, соответствующую цветности стандартного источника освещения Д65, метамер которого использовался нами как постоянный нейтральный фон. Этот фон у наблюдателя, адаптированного к нему, не должен вызывать ощущений ни синих, ни желтых оттенков. Вдоль этой линии мы выбрали пять точек с координатами L/(L+M) 0,60, 0,62, 0,64, 0,6552 (Д65) и 0,68. Через эти пять точек были проведены пять линий ортогонально к линии Д65–576 нм – эти линии имеют наклон +45 градусов. На каждой из этих линий было выбрано семь точек в качестве референтных стимулов (рис. 1, правая панель). Пороги различения цветов, таким образом, были измерены в 35 точках диаграммы цветности МакЛаода-Бойнтона.
Расстояние от наблюдателя до экрана монитора составило 57 см. Стимулы предъявляли на постоянном фоне, метамере источника Д65, который оставался также на экране между предъявлениями.
На рис. 2 (левая панель) приведен пример расположения двух половин диска, цвета которых требовалось различать. Для фиксации четыре маленькие черные точки были расположены в вершинах ромба таким образом, чтобы диск находился в середине этого ромба. Эти точки были постоянно на экране – во время предъявления стимулов и между ними во время ожидания ответа наблюдателя. Длительность стимулов составляла 100 мсек.
Процедура. Пороги цветоразличения измеряли методом пространственного вынужденного выбора. В каждой отдельной серии измеряли различение цветов вдоль одной из пяти линий, идущих по углом +45 градусов к абсциссе и показанных на рис. 2 (правая панель). Референтные стимулы наблюдателям не показывали, а в каждом предъявлении на экране присутствовали только два тестовых стимула, которые находились на той же самой линии на диаграмме цветностей по разные стороны от тестового стимула. Разница в цвете между тестовыми стимулами и референтным стимулом увеличивалась или уменьшалась в зависимости от правильности ответа. Наблюдателя просили нажать правую или левую кнопку на пульте в зависимости от того, какая половина диска казалась зеленоватой (или красноватой) по сравнению со второй половиной диска.
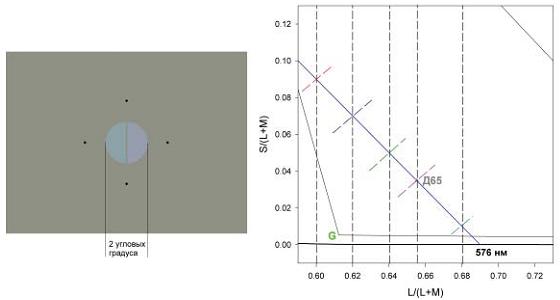
Рис. 2. Левая панель. Показана центральная часть экрана монитора со стимулом (референтный стимул соответствует точке пересечения линии под углом +45 с сине-желтой линией в точке Д65). Правая панель. Часть диаграммы цветностей с измененным таким образом масштабом, чтобы сине-желтая линия проходила под углом -45 градусов к абсциссе (часть диаграммы, приведенной на рис. 1, правая панель). Линия, проходящая под углом -45 – предварительная оценка сине-желтой линии, которая проходит через Д65 и точку чистого желтого, 576 нм. На этой линии лежат цвета, которые для среднего наблюдателя не содержат ни красных, ни зеленых оттенков. Короткие штриховые линии, ортогональные к сине-желтой линии и проходящие под углом +45 градусов к абсциссе, показывают линии, вдоль которых находились референтные и тестовые стимулы и вдоль которых были измерены пороги цветоразличения. Вертикальные штриховые линии показывают точки, в которых линии, проходящие под углом +45 градусов, пересекают сине-желтую линию.
Так как субъективная оценка цвета сильно зависит от положения стимула на наклонной линии и поскольку мы хотели помешать наблюдателям использовать центральную тенденцию (которая привела бы к минимальным порогам для референтного стимула, находящегося в середине диапазона цветов всех семи референтных стимулов каждой серии), эксперимент состоял из нескольких блоков. Порог в каждом блоке измеряли отдельно для каждого референтного стимула, но пороги для всех референтных стимулов, находящихся на одной ортогональной линии, измеряли в одной серии. После ответа наблюдателя звуковой сигнал указывал на правильный или неправильный ответ. Пороги цветоразличения измеряли с помощью процедуры «лестница», которая прекращалась после 15 изменений направлений ответа. Значения последних 10 перегибов усредняли для получения порога. Расстояние между референтным и тестовым стимулами уменьшалось после трех последовательных правильных ответов и увеличивалось после одного неправильного, таким образом, полученный нами порог соответствовал 79,4% правильных ответов на психометрической функции (Wetherill, Levitt, 1965). Цветности референтных и тестовых стимулов сначала задавались в единицах абсциссы (т. е. их L/(L+M) координаты, или l), а затем соответствующие S/(L+M) координаты вычислялись таким образом, чтобы тестовые стимулы лежали на одной и той же прямой. В каждом предъявлении у одного из тестовых стимулов была координата l, а у другого – l. l равнялась l-координате референтного стимула, умноженной на фактор . , а lt2 равнялась l-координате референтного стимула, разделенной на фактор ., где . всегда >1,0. После трех правильных ответов эта величина уменьшалась на 10 %, а после одного неправильного ответа она увеличивалась на 10%, оставаясь всегда больше 1,0.
Пороги различения для семи референтных стимулов вдоль каждой из пяти линий измеряли в одной экспериментальной серии, но каждую из пяти линий использовали в разных сессиях, которые могли проходить в разные экспериментальные дни. Каждая серия повторялась пять раз. Порядок серий и порядок референтных стимулов внутри каждой серии были случайными. У наивных наблюдателей измеряли пороги цветоразличения только вдоль одной из пяти линий, которая пересекала линию Д65–576 нм в точке с координатой L/(L+M)=0,62.
Линия Д65–576 нм была нами взята как предварительная оценка сине-желтой линии, полученная в других аналогичных исследованиях. Для наблюдателей ДДМ и МВД были получены индивидуальные эмпирические данные для построения индивидуальной границы между категориями. В перемежающихся экспериментальных сериях мы измеряли положение точки перехода от красноватых к зеленоватым оттенкам, в то время как цветности стимулов изменялись вдоль тех же пяти линий с наклоном +45 градусов, вдоль которых измеряли пороги цветоразличения. В этой серии стимулом был целый диск того же диаметра, что и в случае измерения порогов цветоразличения, предъявляемый также на 100 мсек. Наблюдателей просили путем нажатия тех же кнопок на пульте указать, какой оттенок присутствует в цвете диска – красноватый или зеленоватый. И для англоговорящих, и для русскоговорящих наблюдателей красный и зеленый цвета являются однозначными. Чтобы уменьшить влияние повторения одного и того же или очень близких стимулов, мы использовали одновременно четыре случайным образом перемежающиеся «лестницы», две из которых начинались с предъявления заведомо зеленых оттенков, а две – заведомо красных оттенков (Jordan, Mollon, 1995). Как и в случае цветоразличения, каждая «лестница» завершалась после 15 изменений категорий ответов, а последние 10 точек перегиба усреднялись. Затем проводилось усреднение четырех таким образом полученных значений, и этот результат рассматривался как полученная в данной серии оценка положения цвета, который не содержит ни красного, ни зеленого оттенков. Эти серии также повторяли пять раз для каждого наблюдателя; порядок измерения точки перехода для каждой из пяти точек пересечения с линией Д65–576 нм был случайным в каждой серии.
Эксперимент 1 а. Результаты
На рис. 3 приведены положения пороговых значений тестовых стимулов, нанесенные на диаграмму цветности МакЛаода-Бойнтона. Каждая пара точек соединена короткой линией и представляет собой положение двух тестовых стимулов, которые наблюдатель различает в 79,4% предъявлений. Вдоль каждой из линий, идущих под углом +45 градусов к абсциссе, наблюдается изменение расстояния между двумя тестовыми стимулами: наименьшие расстояния между ними (и, следовательно, лучшее различение) локализованы близко к середине измерения диапазона тестовых стимулов.
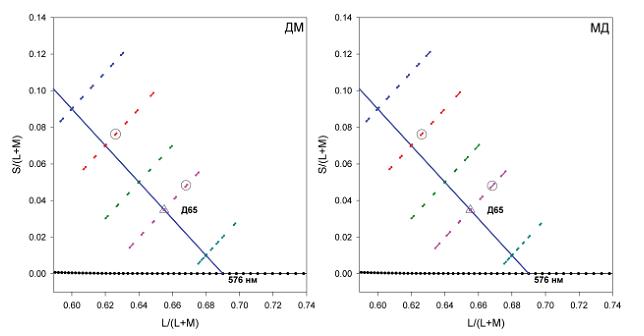
Рис. 3. Малая часть масштабированной диаграммы цветностей МакЛаода-Бойнтона. Каждая пара точек, соединенная короткой линией, показывает положение двух тестовых стимулов в момент различения в 79,4% случаев, т. е. в точке порогового различения, для двух наблюдателей, поэтому длина штриха отражает величину порога. Следует отметить, что порог для референтного стимула с координатами 0,6262, 0,0763 (штрих, заключенный в кружок на красной линии) меньше, чем порог для референтного стимула с координатами 0,6683, 0,048 (штрих, заключенный в кружок на розовой линии), хотя последний находится намного ближе к Д65 как в единицах L/(L+M), так и в единицах S/(L+M)
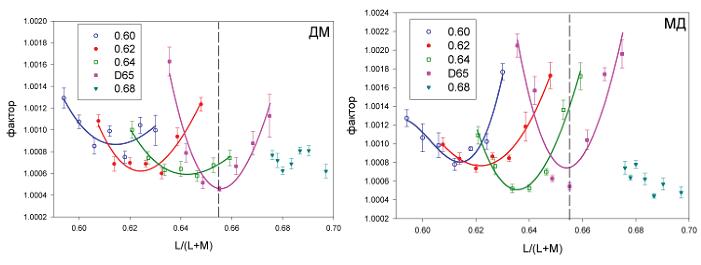
Рис. 4. Пороги в единицах . в зависимости от L/(L+M) координаты референтного стимула. Каждый набор точек (обозначенный разными символами) соответствует одной из линий, идущих под углом +45 градусов и перпендикулярных сине-желтой линии (см. рис. 2, правая панель). На легенде приведены значения L/(L+M), в которых соответствующая линия пересекает исходную сине-желтую линию. Ордината представляет фактор . в единицах L/(L+M), при котором два тестовых стимула различаются с точностью 79,4% (см. Методику). Сплошные линии представляют аппроксимацию экспериментальных точек обратными полиномами третьего порядка; форма кривых была выбрана условно для оценки минимальных порогов цветоразличения, которые приведены на рис. 5, и не имеет никакой теоретической значимости.
Масштаб изменения порогов цветоразличения в разных участках диаграммы цветностей можно более наглядно увидеть на рис. 4, где приведены пороговые значения фактора . для пяти серий. Абсцисса пороговых значений факторов – координата L/(L+M), или l-координата соответствующего референтного стимула. Вертикальные пунктирные линии показывают l-координату нейтрального фона метамера Д65, который мы использовали в эксперименте. Ордината представляет значение фактора ., на который каждая из половин диска отличалась от соответствующей l-координаты референтного стимула (см. Методику). Приведенные на рис. 4 значения факторов составляют примерно половину соответствующих традиционных отношений Вебера. Сплошные линии – аппроксимация семи экспериментальных значений факторов, полученных вдоль одной из пяти линий. Для аппроксимации мы применили обратные полиномы третьей степени. Эти кривые использовались только для нахождения минимального фактора в каждой серии данных (см. рис. 5, незаполненные кружки), но они не несут никакого теоретического смысла.
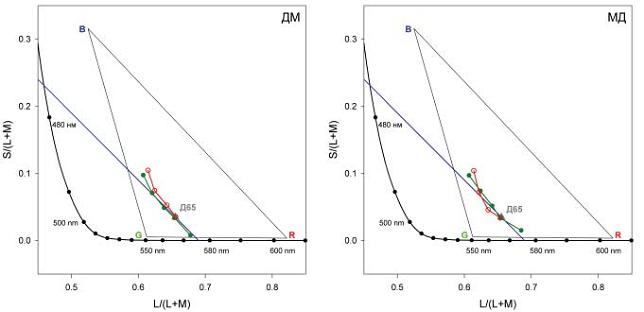
Рис. 5. Часть масштабированной диаграммы цветностей МакЛаода-Бойнтона, на которой сравниваются положения минимальных порогов цветоразличения и точек перехода от красных к зеленым оттенкам. Заполненные кружки представляют цвета, которые наблюдатели оценивают как не содержащие ни красных, ни зеленых оттенков. Незаполненные кружки представляют положения минимальных порогов, полученных в результате аппроксимации полиномами экспериментальных измерений по методике вынужденного выбора (см. рис. 4)
Результаты двух наблюдателей схожи между собой: в случае четырех из пяти линий, которые пересекают сине-желтую линию в точке, равной L/(L+M) значению цветности Д65 и меньших значений L/(L+M), значения факторов имеют минимальные значения, а абсолютные величины . находятся между 1,0005 и 1,001. В случае последней, пятой серии (самый правый набор данных, который соответствует желтым цветам) выраженный минимум отсутствует, – аналогичный результат мы получили и в случае парафовеальных данных (Danilova, Mollon, 2010).
На рис. 5 приведен участок диаграммы цветностей МакЛаода-Бойнтона, где нанесены положения переходных точек от красноватых к зеленоватым цветам (сплошные кружки), определенные из феноменологических серий (см. Методику). В единицах l-координаты средние величины стандартных ошибок средних значений точек перехода составляли 0,0003 (МД) и 0,0008 (ДМ): эти величины получены как результат усреднения серий, проведенных в разные экспериментальные дни, и показывают, что наблюдатели могут производить феноменологические оценки с высокой степенью повторяемости результата.
Эксперимент 1б. Результаты
На рис. 6 приведены результаты эксперимента с участием шести наивных наблюдателей, у которых пороги цветоразличения измеряли только вдоль одной из пяти линий, которая пересекает линию Д65–576 нм в точке L/(L+M)=0.62. Каждый наблюдатель выполнил шесть повторений серии, из которых пять последних были усреднены. На рисунке приведены усредненные данные всех шести наблюдателей, а ошибки на графике представляют стандартные ошибки среднего. Дисперсионный анализ подтверждает достоверность изменения порогов различения в зависимости от положения референтного стимула на линии, проходящей под углом +45 градусов к абсциссе и пересекающей ось Д65–576 нм в точке L/ (L+M)=0,62: F[Beer, 2006]=16,63, p<0,0001.
Эксперимент 1 а,б. Обсуждение
При использовании традиционного фовеального стимула (диска размером 2 угловых градуса, разделенного на две части) мы обнаружили наличие ограниченного участка на диаграмме цветности, где наблюдаются более низкие пороги различения цветов. Этот результат согласуется с нашими данными, полученными в парафовеа (Danilova, Mollon, 2010). Данное явление встречается как у опытных наблюдателей, так и у наивных, которые не знали целей эксперимента.
Если бы положения минимальных порогов цветоразличения совпадали с точкой на диаграмме цветности, где нейронный канал, определяющий отношение возбуждений L-и M-колбочек (см. Введение), находится в равновесии, тогда минимум не должен зависеть от величины сигнала, поступающего от S-колбочек, и всегда должен находиться на вертикальной линии, соответствующей одному и тому же значению L/(L+M). Другими словами, положения минимумов должны иметь всегда одно и то же значение на абсциссе для всех серий данных на рис. 4. Из рисунка же видно, что минимумы имеют разные значения абсциссы. С другой стороны, ординаты минимумов также не соответствуют одному и тому же значению S/(L+M). Вместо этого при увеличении значений ординат минимальных порогов цветоразличения значения абсцисс уменьшаются, т.е. при увеличении сигнала, поступающего в S-колбочки, минимальные пороги цветоразличения наблюдаются при меньших относительных возбуждениях L- и M-колбочек.
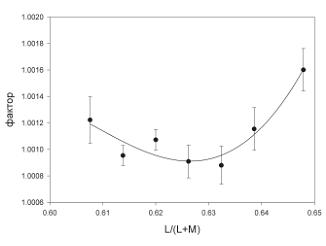
Рис. 6. Среднее значение порогов, полученное на шести наивных наблюдателях (эксперимент 1 б). Ордината графика представляет собой фактор α в единицах L/(L+M), на который два тестовых стимула отличаются от референтного (см. Методику). Семь референтных стимулов находятся на кривой, проходящей под углом +45 градусов к абсциссе, и пересекают сине-желтую линию Д65–576 нм в точке с координатой L/(L+M)=0.62. Погрешности на графике представляют среднеквадратичные ошибки
среднего шести наблюдателей. Сплошная линия – аппроксимация данных обратными полиномами третьего порядка, которые не имеют теоретического значения
Рис. 5 показывает, что для двух наблюдателей минимумы цветоразличения находятся около сине-желтой линии, которая определена для тех же наблюдателей из их феноменологических оценок. Другими словами, минимальные пороги цветоразличения расположены вблизи границы между двумя категориями цветов, т.е. вблизи границы между красноватыми и зеленоватыми оттенками цветов.
Эксперимент 2. Контрольный эксперимент
Является ли обнаруженный нами эффект наличия минимальных порогов цветоразличения на границе категорий действительно сенсорным эффектом, или мы имеем дело с неопределенностью в упорядочении двух цветов, принадлежащих одной категории, и определении того цвета, у которого зеленый оттенок более выражен?
Для ответа на этот вопрос мы предложили наблюдателям оценивать только наличие или отсутствие цветовых различий без определения направления различий. Если в эксперименте 1 наблюдателя просили сказать, какая из половин диска более зеленая (или более красная), то теперь мы спрашиваем только, одинаковые или разные по цвету эти две половины. Традиционная процедура «одинаковый–разный», где предъявляются только два стимула и необходимо сказать, одинаковые они или разные, в данной ситуации не очень подходит, поскольку критерий может меняться в случае разных цветов. Для этого эксперимента мы применили вариант процедуры «одинаковый–разный», но наблюдатель был поставлен в условия вынужденного выбора, – аналогичная процедура была использована в экспериментах по цветоразличению Моллоном и Кавониусом (Mollon, Cavonius, 1987). Каждое предъявление состояло из двух временных интервалов, каждый из которых содержал диск, разделенный на две части. В одном интервале (он мог быть и первым, и вторым) две половины диска были одинаковые по цвету, а во втором интервале они отличались. Задачей наблюдателя в этом эксперименте было указать, в каком интервале – первом или втором – находились две разные по цвету половины диска.
Эксперимент 2. Методика
Наблюдатели. В эксперименте 2 принимали участие два автора статьи (МВД и ДДМ), а также три наивных наблюдателя, не осведомленных о целях эксперимента. Наивные наблюдатели не имели нарушений цветового зрения (проверены с использованием Cambridge Colour Test), их родной язык был английский, они принимали участие также и в эксперименте 1а.
Аппаратура и стимулы. В эксперименте применялась та же аппаратура, что и в экспериментах 1 а и 1 б. Стимулы также были похожи, только вместо одного временного интервала каждое предъявление состояло из двух временных интервалов длительностью 100 мсек, разделенных 500 мсек. В одном временном интервале две половины диска были одинаковые по цветности, а в другом интервале они различались. Так же как и в эксперименте 1, средняя яркость дисков была на 30% выше, чем яркость фона, которая составляла 10 кд/м2 в единицах яркости МКО1931. Яркость каждой половины диска независимо варьировали в пределах ± 5%.
Процедура. Измерения порогов цветоразличения были проведены только вдоль одной из пяти линий, проходящих под углом +45 градусов к абсциссе и использовавшихся в эксперименте 1: эта линия пересекает Д65–576 нм ось в точке с координатой L/(L+M)=0,62. Как и в эксперименте 1, пороги различения были измерены для каждого из семи референтных стимулов вдоль выбранной линии. Предъявлялись не сами референтные стимулы, а только два тестовых стимула с координатами L/(L+M) больше и меньше референтного. В зависимости от ответа наблюдателя расстояние на диаграмме цветностей между тестовыми стимулами уменьшали или увеличивали по тому же правилу «лестницы», что и в эксперименте 1. Пороги различения для разных референтных стимулов измеряли в одной серии в случайном порядке. Серии повторяли не менее пяти раз, но большинство наблюдателей выполнили 8–10 серий, и, таким образом, каждая точка для каждого наблюдателя является средним значением по меньшей мере пяти повторений.
Эксперимент 2. Результаты и обсуждение
На рис. 7 изображена кривая, показывающая средние значения порогов цветоразличения пяти наблюдателей, принимавших участие в эксперименте 2. Ошибки среднего, приведенные на графике, представляют среднеквадратичные ошибки среднего пяти наблюдателей. Дисперсионный анализ подтвердил, что порог достоверно меняется в зависимости от положения референтного стимула: F[Beer, 2006]=2,73, p=0,036.
Таким образом, эффект, который был получен в эксперименте 1, сохраняется при изменении задачи, поставленной перед наблюдателем. Даже если не требуется определить присутствие того или иного оттенка в стимуле, существуют цвета, пороги различения которых минимальны. Этот результат предполагает, что полученный нами в эксперименте 1 эффект не связан с соотношением категории цвета и его называнием.
Общее обсуждение
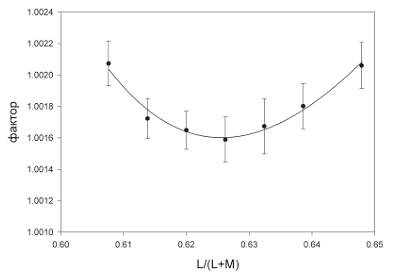
Рис. 7. Результаты эксперимента 2, в котором определяли пороги цветоразличения, используя методику вынужденного выбора в случае оценки «одинаковый–разный». Точки представляют среднее значение для пяти наблюдателей. Ордината графика – фактор α в единицах L/(L+M), на который два тестовых стимула отличаются от референтного в интервале, где предъявлялись разные стимулы (см. Методику). Семь референтных стимулов находятся на кривой, проходящей под углом +45 градусов к абсциссе, и пересекают сине-желтую линию Д65–576 нм в точке с координатой L/(L+M)=0,62. Погрешности на графике представляют среднеквадратичные ошибки среднего пяти наблюдателей. Сплошная линия – аппроксимация данных обратными полиномами третьего порядка, форма кривых не имеет теоретического значения
Общее обсуждение
Нейронные каналы и когнитивные цветовые категории
В области диаграммы цветностей вдоль сине-желтой линии, где представлены цвета средней насыщенности тонов от синих (с левой стороны от точки Д65 диаграммы) до беловатых (в области Д65) и желтоватых (справа от Д65), находится область низких порогов различения, т. е. улучшен- ного цветоразличения. Эта область расположена с отрицательным наклоном на диаграмме приблизительно вдоль сине-желтой линии и поэтому не соответствует точке равновесия парвоцеллюлярного канала, который определяет отношение возбуждений L- и M-колбочек: если различение цветов в наших экспериментах зависит от работы этого канала, то тогда минимальные пороги должны наблюдаться всегда при одном и том же значении L/(L+M). Это значение L/(L+M) должно соответствовать значению L/(L+M) нашего постоянного фона – Д65, так как нейронные каналы наиболее чувствительны к малым отклонениям от цвета, к которому адаптирована зрительная система (De Valois et al., 1967; Раутиан, Соловьева, 1954). Если же различение цветов в нашем эксперименте зависит от работы канала, который получает вход от малых бистратифицированных ганглиозных клеток сетчатки, то следует ожидать минимальные пороги в области цветов с координатой S/(L+M) фона Д65.
Вместе с тем, область обнаруженных нами минимальных порогов соответствует (хотя и не совсем точно) границе между субъективными категориями цветов с оттенками красного, с одной стороны, и зеленого – с другой. Эта граница представляет собой линию, проходящую через чистый синий и чистый желтый. В качестве первоначальной оценки этой линии мы выбрали линию, которая проходит через чистый желтый (576 нм) и Д65. Эта же линия проходит через точку 478 нм: согласно литературным данным, эта точка лежит в пределах чистых синих цветов для разных наблюдателей. В наших экспериментах для наблюдателей МВД и ДДМ в сериях, случайным образом перемешанных с сериями по измерению порогов цветоразличения, мы также определили индивидуальные феноменологические границы между зеленоватыми и красноватыми цветами (рис. 5). Эта граница проходит примерно параллельно положению минимальных порогов. В психофизических экспериментах не следует ожидать точного совпадения субъективных оценок и количественного измерения порогов различения. С одной стороны, определение минимальных порогов в нашем случае зависит от приближения произвольными функциями дискретных точек, полученных в эксперименте. С другой стороны, оценка положения чистых цветов (хоть и хорошо воспроизводимая в разные экспериментальные дни) зависит от субъективного критерия каждого наблюдателя, в частности, от применения определенной стратегии в тех случаях, когда отнесение бинарных цветов к одной из категорий в области, близкой к переходу, затруднено. При постановке задачи кажется достаточно очевидным просить наблюдателя нажать определенную кнопку, если диск кажется красноватым, и другую – если он содержит зеленоватые оттенки. Однако при длительностях предъявления 100 мсек и цветах, близких к границе перехода, появляющийся на короткое время диск никогда не кажется однородным: наблюдатели сообщали, что часто разные участки диска казались зеленоватыми, а другие части в то же время казались красноватыми.
Красно-зеленый канал
До сих пор из обсуждения результатов наших экспериментов может показаться, что на пороги различения существенно влияет граница между категориями бинарных цветов (красноватые и зеленоватые), которая выявляется на когнитивном, центральном, уровне. Однако возникает вопрос, который уже поднимался во Введении: существует ли нейронный канал, функционирование которого смогло бы объяснить и наличие минимальных порогов цветоразличения, и наличие перехода от бинарных красноватых цветов к бинарным зеленоватым цветам? Такой гипотетический канал должен отличаться от каналов, соответствующих основным осям диаграммы цветностей МакЛаода-Бойнтона (см. Введение). Его вход должен образовываться сигналами противоположного знака от M-колбочек и от комбинации L- и S-колбочек. В исследовательской литературе встречаются сообщения о существовании нейронов, для которых входы S- и L-колбочек синергичны (De Monasterio et al., 1975; Valberg et al., 1986). Если предположить существование канала, получающего противоположные входы от M-колбочек, с одной стороны, и от комбинации L- и S-колбочек – с другой, то тогда цвета, при которых пороги различения будут минимальны, должны соответствовать цветам, которые приводят этот предполагаемый канал в равновесие. Такие цвета должны обеспечивать постоянное отношение сигналов от M-колбочек к комбинации сигналов от S- и L-колбочек. Если бы такой канал существовал в зрительной системе, то его работа естественным образом объяснила бы существование перехода от красных к зеленым цветам и никаких специальных преобразований на когнитивных уровнях не требовалось бы для объяснения наличия границы между этими категориями.
Предыдущие исследования
Предлагаемые нами данные о наличии такого канала согласуются с известными результатами измерения эллипсов различения. Бойтон и другие (Boynton, et al., 1983) определяли эллипсы различения и показали существование одного эллипса, центр которого находился в области беловато-синеватых цветов, где пороги различения были минимальны в направлении +45 градусов к абсциссе. Схожие результаты были получены в других работах (Beer et al., 2006), где просили наблюдателей модифицировать цвет диска на черном фоне так, чтобы диск казался белым. Все наблюдатели использовали очень узкий диапазон оттенков белого в направлении красный–зеленый, но диапазон их установки вдоль оси синий–желтый был гораздо более широкий, что указывает на более точное различение отклонений от белого цвета в направлении +45 градусов на диаграмме цветностей. Другие авторы (Krauskopf, Gegenfurtner, 1992) приводят описание двух эллипсов, которые расположены на диаграмме цветностей с разных сторон точки, соответствующей адаптирующему белому цвету. В обоих случаях эллипсы расположены так, что их длинная ось направлена вдоль сине-желтой оси в сторону белого фона, т. е. пороги различения минимальны в направлении +45 градусов к абсциссе.
Выводы
Как в парафовеа, так и в фовеа существует ряд цветов, около которых минимальные пороги цветоразличения могут быть измерены с помощью методики вынужденного выбора (эксперимент 1). Оптимальное цветоразличение – явление сенсорное, а не результат неправильной оценки наличия второго цвета (красноватого или зеленоватого) в цвете одной и той же категории (эксперимент 2). Положение точек оптимального цветоразличения примерно соответствует положению границы между красными и зелеными оттенками, однако наличие такого участка невозможно объяснить двумя оппонентными цветовыми каналами, на существование которых в зрительной системе указывают данные электрофизиологических и психофизических экспериментов. Наличие минимальных порогов цветоразличения и положение субъективной границы между красными и зелеными цветами можно объяснить, предположив существование нейронного канала, который получает входы противоположного знака от M-колбочек и от комбинации S- и L-колбочек. Однако наличие такого канала не объясняет, почему область оптимального цветоразличения ограничена частью диаграммы цветности в районе точки белого адаптирующего фона и не выявляется при удалении от фона в обе стороны по направлению к чистым цветам – синему и желтому.