Современный системный подход указывает на то обстоятельство, что для целостного понимания всякого сложноорганизованного психического феномена необходимо рассматривать его в процессе развития (Ломов, 1999), причем как онтогенетического, так и филогенетического. Данный тезис, в частности, имеет отношение и к такой структуре человеческой психики, как Я-концепция. Вопрос специфики восприятия себя человеком, а также его развития в онтогенезе детально изучен в работах зарубежных и отечественных психологов. Однако филогенетические предпосылки этого феномена практически не привлекали внимания ученых за исключением представителей отдельных видов, эволюционно наиболее близких к человеку. Так широкоизвестны эксперименты Г. Гэллапа (Gallup, 1970), направленные на выявление способности к самоузнаванию в зеркале у шимпанзе. Позже подобные эксперименты проводились на других видах позвоночных (Зорина, Полетаева, 2003), и недавно были получены данные, позволяющие говорить о наличии такой способности у слонов (Plotnik et al., 2006), дельфинов (Reiss, Marino, 2001) и врановых (Prior et al., 2008). Отметим, что методики, использованные в данных экспериментах, были однотипными.
На наш взгляд, перспективным представляется выявление антропогенетически значимых черт и предпосылок становления человеческой Я-концепции у животных, относящихся к наиболее ранним стадиям эволюции психики. В настоящей статье описывается эксперимент, целью которого являлось изучение специфики самоотражения у представителей низшего уровня перцептивной стадии развития психики (по периодизации Г. Г. Филипповой, 2004).
Под самоотражением мы понимаем процесс и результат отражения субъектом своей внутренней объективной реальности – характеристик своего организма; в контексте данного исследования речь будет вестись о размерах и границах собственного тела. Ранее с опорой на схему перцептивного образа В. А. Барабанщикова (2002) нами была разработана концептуальная модель, согласно которой самоотражение рассматривалось системно, в контексте своей взаимосвязи с отражением внешней среды, как двух элементов единого образа мира (Хватов, 2009, 2010 a, 2010 б). Движущей силой развития данной системы является противоречие между формой и содержанием отражения (Барабанщиков, 2002), приводящее далее к возникновению противоречий между самоотражением и отражением внешнего мира, проявляющихся сначала в межсистемных, а затем во внутрисистемных связях, что соответствует общему представлению о движущих силах процесса эволюции психики А. Н. Леонтьева (1981).
Проанализировав с помощью данной модели особенности самоотражения у животных, находящихся на разных стадиях развития психики, с опорой на общее представление об эволюции психического отражения (Леонтьев, 1981; Фабри, 2004; Филиппова, 2004), мы выделили магистральную линию развития самоотражения в филогенезе. Она заключается в постепенной дифференциации внутри психического отражения – дифференциации между отражением субъектом внешнего мира и самого себя. Данное положение согласуется с фундаментальным дифференционно-интеграционным законом развития (Чуприкова, 2007). Было также показано, что в развитии самоотражения в филогенезе наличествуют моменты, аналогичные (но не тождественные) таковым в процессе развития самоотражения в онтогенезе человека.
Соответственно, применительно к цели настоящего исследования была выдвинута гипотеза о том, что представители низшего уровня перцептивной стадии развития психики способны отражать пространственные характеристики своего тела (границы своего тела) в виде переживаний, возникающих в процессе взаимодействия с объектами внешнего мира. Однако у этих животных самоотражение и отражение внешнего мира все еще тесно связаны в структуре единого образа и не существуют отдельно друг от друга.
Методика экспериментального исследования
Характеристика испытуемых. В качестве испытуемых представителей низшего уровня перцептивной стадии развития психики для настоящего эксперимента были избраны имаго самцы тараканов вида Periplaneta americana (американский таракан) – 90 особей. Данный выбор объясняется, во-первых, тем, что по своим физическим характеристикам тараканы в наибольшей степени подходят к специфике нашего исследования (The American cockroach, 1982), во-вторых, эти животные наиболее доступны. Для определения того, способны ли данные насекомые отражать границы собственного тела, важным являлось то обстоятельство, что в построении образа мира у тараканов одна из главнейших ролей отводится кинестетическому анализатору при относительно незначительной роли зрения, кроме того, физиология и психика этих животных сравнительно хорошо изучены (The American cockroach, 1982).
Оборудование и план эксперимента. Эксперимент проводился по методике лабиринта. Лабиринт располагался в коробке размерами 300 на 600 мм и высотой стенки 40 мм, накрытой сверху стеклянной крышкой (рис. 1). В качестве отрицательного подкрепления использовался яркий свет, покрывавший большую часть лабиринта. Тараканы обладают выраженным отрицательным фототаксисом (The American cockroach, 1982), поэтому стараются как можно быстрее скрыться с освещенных участков и попасть в затемненную область лабиринта – «выход». Между входом и выходом располагалась перегородка, в которой имелись два отверстия полукруглой формы.
Ставилась задача выявить, способны ли испытуемые менять свое поведение сообразно изменению размеров и границ собственного тела. Размеры тела изменялись с помощью пенопластового куба со стороной 15 мм, прикреплявшегося на переднюю часть спины насекомого.
В эксперименте принимали участие три выборки (см. табл. 1): в первой варьировались размеры тела насекомого; во второй – размеры отверстий в лабиринте; в третьей – обе эти переменные. Эксперимент проводился в три этапа (табл. 1).
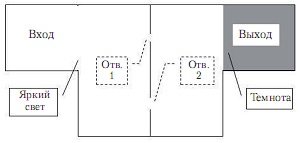
Рис. 1. План лабиринта
На первом этапе у всех испытуемых формировалось научение прохождения лабиринта через отверстие № 1, т. е. кратчайшим путем. На втором этапе определялось, смогут ли испытуемые первой выборки сформировать новое научение при условии: их увеличенное тело не проникает в среднее по размеру отверстие (полукруг с диаметром 15 мм); у испытуемых второй выборки, наоборот, уменьшалось отверстие № 1 до непроходимости (полукруг с диаметром 3 мм); у испытуемых третьей выборки отверстие № 1 изначально было большим (полукруг с диаметром 25 мм), и в него можно было пройти даже с увеличенным телом. На третьем этапе определялось то, насколько быстро животные переучиваются в случае варьирования независимых переменных.
Таблица 1. Схема эксперимента
|
Этапы |
Выборка № 1 (30 тараканов) |
Выборка № 2 (30 тараканов) |
Выборка № 3 (30 тараканов) |
|
Этап № 1 50 проб |
Форма тела испытуемых – исходная Отверстие № 1 – среднее Отверстие № 2 – большое |
Форма тела испытуемых – исходная Отверстие № 1 – среднее Отверстие № 2 – большое |
Форма тела испытуемых – исходная Отверстие № 1 – большое Отверстие № 2 – большое |
|
Этап № 2 50 проб |
Форма тела испытуемых – измененная Отверстие № 1 – среднее Отверстие № 2 – большое |
Форма тела испытуемых – исходная Отверстие № 1 – маленькое Отверстие № 2 – большое |
Форма тела испытуемых – измененная Отверстие № 1 – большое Отверстие № 2 – большое |
|
Этап № 3 50 проб |
Форма тела испытуемых – измененная Отверстие № 1 – большое Отверстие № 2 – среднее |
Форма тела испытуемых – исходная Отверстие № 1 – большое Отверстие № 2 – маленькое |
Форма тела испытуемых – измененная Отверстие № 1 – среднее Отверстие № 2 – большое |
Зависимой переменной, фиксируемой в ходе эксперимента, было количество проникновений и попыток проникновения в отверстия (суммировалось количество проникновений у всех испытуемых отдельной выборки в каждой пробе). Поведенческим критерием, подтверждающим гипотезу исследования о способности тараканов отражать (учитывать) границы собственного тела, являлось изменение способа прохождения лабиринта испытуемыми выборки № 1 на втором и третьем этапах эксперимента, вызванное изменением размеров их тела. Также сравнивались показатели испытуемых трех выборок между собой для определения того, насколько отличается их поведение в зависимости от того, меняются ли размеры их тела или размеры отверстий лабиринта.
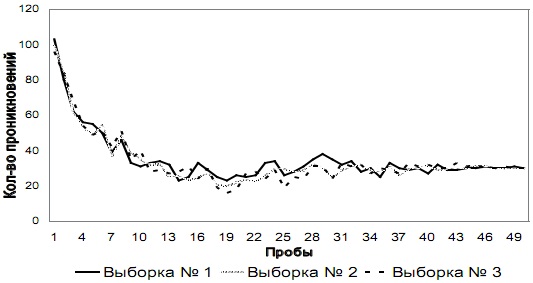
Рис. 2 а. Количество проникновений и попыток проникновения в отверстие №1 на этапе № 1 у испытуемых трех выборок
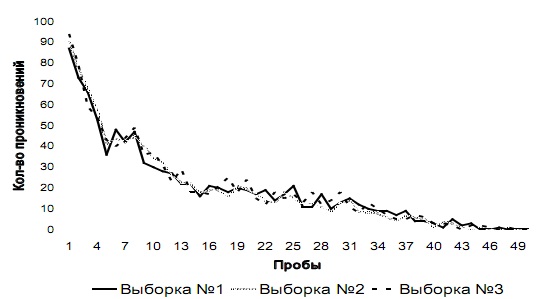
Рис. 2 б. Количество проникновений и попыток проникновения в отверстие № 2 на этапе № 1 у испытуемых трех выборок
Результаты исследования
Этап № 1. Общая динамика изменения количества проникновений в отверстия № 1 и № 2 у испытуемых трех выборок на этапе № 1 показана на рис. 2 а и 2 б соответственно.
Как видно из графиков, количество проникновений в оба отверстия резко снижается к середине этапа у испытуемых всех трех выборок. На последних пяти пробах этапа суммарное количество проникновений в отверстие № 2 за каждую пробу составляет в среднем 0 (SD=0,45), 100 ![]() а в отверстие № 1 составляет 30 за пробу (SD=0,45).
а в отверстие № 1 составляет 30 за пробу (SD=0,45).
Качественная характеристика поведения животных. На первых пробах испытуемые всех трех выборок демонстрировали высокую локомоторную активность. Насекомые быстро бегали по полу, стенам и потолку лабиринта, пытаясь забиться в любой угол. Многократно проникали в отверстия, часто бегая кругами, проходя из одного отверстия в другое.
Траектория движения сложная: покрывает практически всю площадь лабиринта, включая стены. Особенно активно ощупывались (лапками и антеннами) места соединения пола со стенами и углы. В светлой части лабиринта проводят достаточно продолжительное время. После проникновения в затемненную часть лабиринта насекомые резко снижали локомоторную активность, прижавшись к стене или к углу, и более не покидали этого отсека. На последних пробах испытуемые сразу после попадания в лабиринт направляются к отверстию № 1 и, минуя его, движутся к затемненному отсеку лабиринта, где демонстрируют поведение, аналогичное тому, что проявлялось в данном отсеке на первых пробах. Траектория движения простая – в подавляющем большинстве случаев представляет собой прямую линию, ведущую от входа к выходу через отверстие № 1. В светлой части лабиринта испытуемые находятся всего несколько секунд.
С помощью критерия χ2 с поправкой на непрерывность, так как df=1 (Сидоренко, 2010), была выполнена проверка нулевой гипотезы об отсутствии различий между эмпирическим распределением количества проникновений в отверстия № 1 и № 2 на каждой пробе этапа № 1 у испытуемых трех выборок и равномерным распределением. В табл. 2 приведены данные по 1, 10, 20, 30, 40 и 50 пробам. Как видно из табл. 2, начиная с 40-й пробы нулевая гипотеза об отсутствии различий между эмпирическим и равномерным распределением должна быть отвергнута, т.е. количество проникновений в отверстие № 1 достоверно преобладает над количеством проникновений в отверстие № 2.
Таблица 2. Значения критерия χ2 при сопоставлении эмпирических распределений количества проникновений в отверстия № 1 и № 2 с равномерным распределением на этапе № 1 у испытуемых трех выборок (достоверные отличия отмечены жирным шрифтом)
|
Пробы |
Выборка №1 (df=1) |
Выборка №2 (df=1) |
Выборка №3 (df=1) |
|||
|
1 |
χ2=1,18 |
p>0,05 |
χ2=0,33 |
p>0,05 |
χ2=0,00 |
p>0,05 |
|
10 |
χ2=0,00 |
p>0,05 |
χ2=0,00 |
p>0,05 |
χ2=0,05 |
p>0,05 |
|
20 |
χ2=0,80 |
p>0,05 |
χ2=0,02 |
p>0,05 |
χ2=0,59 |
p>0,05 |
|
30 |
χ2=9,18 |
p<0,01 |
χ2=3,18 |
p>0,05 |
χ2=0,59 |
p>0,05 |
|
40 |
χ2=17,63 |
p<0,01 |
χ2=27,27 |
p<0,01 |
χ2=22,40 |
p<0,01 |
|
50 |
χ2=28,03 |
p<0,01 |
χ2=28,03 |
p<0,01 |
χ2=28,03 |
p<0,01 |
Также с помощью критерия χ2 мы проверили нулевую гипотезу об отсутствии различий в распределениях количества проникновений в отверстия № 1 и № 2 за 50 проб этапа № 1 (df=49) между испытуемыми трех выборок для сопоставления динамики их научения. Результаты для отверстия № 1: (выборки № 1 и № 2) χ2=24,15, p>0,05; (выборки № 1 и № 3) χ2= 43,20, p>0,05; (выборки № 2 и № 3) χ2= 20,16, p>0,05. Результаты для отверстия № 2: (выборки № 1 и № 2) χ2= 31,10, p>0,05; (выборки №1 и №3) χ2= 49,70, p>0,05; (выборки № 2 и № 3) χ2= 28,38, p>0,05. Таким образом, нулевая гипотеза принимается во всех шести случаях.
Этап № 2. Общая динамика изменения количества проникновений в отверстия № 1 и № 2 у испытуемых трех выборок на этапе № 2 показана на рис. 3 а и 3 б соответственно.
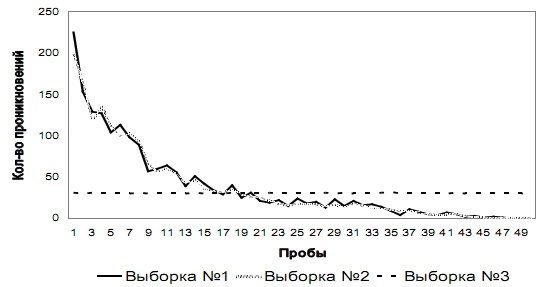
Рис. 3 а. Количество проникновений и попыток проникновения в отверстие № 1 на этапе № 2 у испытуемых трех выборок
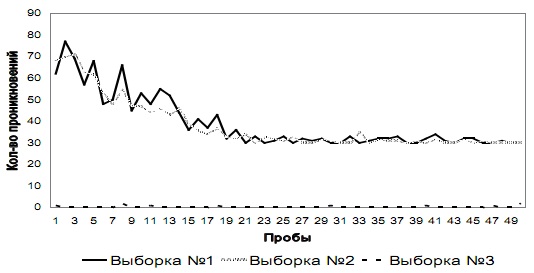
Рис. 3 б. Количество проникновений и попыток проникновения в отверстие № 2 на этапе № 2 у испытуемых трех выборок
Как видно из графиков, у испытуемых выборки № 3 среднее суммарное количество проникновений в оба отверстия на каждой пробе существенно не меняется на всем этапе № 2: среднее количество проникновений на каждой пробе в отверстие № 1 – 30 (SD=0,48); в отверстие № 2 – 0 (SD=0,49). С другой стороны, у испытуемых выборок № 1 и № 2 на первых пробах этапа № 2 возрастает количество проникновений и попыток проникновения в оба отверстия по сравнению с количеством проникновений в данные отверстия у испытуемых этих же выборок на последних пробах этапа № 1. У этих животных количество проникновений в отверстие № 2 снижается до 25-й пробы, а далее в среднем составляет 30 (SD=1,22), а количество проникновений в отверстие № 1 снижается до 45-й пробы и на последних пяти пробах в среднем составляет 0 (SD=0,69).
Качественная характеристика поведения испытуемых выборки № 1 и № 2. На первых пробах этапа № 2 тараканы резко направляются к отверстию №1 и пытаются пройти через него, упираясь в перегородку отверстия. Быстро перебирают лапками в течение нескольких секунд. Затем локомоторная активность животных снова резко возрастает аналогично первым пробам этапа № 1. Насекомые бегают по лабиринту, прижимаясь к углам и стенкам, затем снова пытаются проникнуть в отверстие № 1. Это происходит даже в тех случаях, если они перебегают на противоположную сторону лабиринта через отверстие № 2, – затем они снова возвращаются обратно к отверстию № 1. Траектория движения по лабиринту такая же сложная, как на первых сериях этапа № 1. В светлой части лабиринта насекомые проводят даже больше времени, чем на первых пробах предыдущего этапа, но идут к выходу через другое отверстие. После проникновения в затемненную часть лабиринта ведут себя аналогично предыдущим пробам в этом отсеке. На последних пробах этапа № 2 демонстрируют поведение, аналогичное последним пробам предыдущего этапа. Сразу после попадания в лабиринт направляются к отверстию № 2 и, минуя его, движутся к затемненному отсеку лабиринта, где демонстрируют поведение, аналогичное тому, что проявлялось в данном отсеке на первых пробах. Траектория движения простая, представляет собой прямую линию, ведущую от входа к выходу через отверстие № 2. В светлой части лабиринта испытуемые находятся всего несколько секунд.
Поведение испытуемых выборки № 3 на всех пробах этапа № 2 такое же, как и на последних пробах предыдущего этапа.
С помощью критерия χ2 с поправкой на непрерывность (df=1) была выполнена проверка нулевой гипотезы об отсутствии различий между эмпирическим распределением количества проникновений в отверстия № 1 и № 2 на каждой пробе этапа № 2 у испытуемых трех выборок и равномерным распределением. В табл. 3 приведены данные по 1, 10, 20, 30, 40 и 50 пробам.
Как видно из таблицы, для выборки № 3 нулевая гипотеза отвергается на всех пробах этапа № 2, т. е. количество проникновений в отверстие № 1 достоверно преобладает над количеством проникновений в отверстие № 2. Для выборок № 1 и № 2 на первой пробе значение критерия выше критического, что обусловлено тем, что испытуемые совершали достоверно больше попыток проникновения в отверстие № 1 (в которое они теперь пройти не могли), нежели проникновений в отверстие № 2. Начиная с 10-й пробы, как показано в таблице, у испытуемых этих двух выборок эмпирическое распределение количества проникновений в отверстия № 1 и № 2 не отличается от равномерного (принимается нулевая гипотеза). Однако, начиная с 40-й пробы и до конца этапа, нулевая гипотеза должна быть отвергнута, так как количество проникновений в отверстие № 2 у испытуемых выборок № 1 и № 2 достоверно преобладает над количеством попыток проникновения в отверстие № 1.
Таблица 3. Значения критерия χ2 при сопоставлении эмпирических распределений количества проникновений в отверстия № 1 и № 2 с равномерным распределением на этапе № 2 у испытуемых трех выборок (достоверные отличия отмечены жирным шрифтом)
|
Пробы |
Выборка № 1 (df=1) |
Выборка № 2 (df=1) |
Выборка № 3 (df=1) |
|||
|
1 |
χ2=91,44 |
p<0,01 |
χ2=62,56 |
p<0,01 |
χ2=26,28 |
p<0,01 |
|
10 |
χ2=0,31 |
p>0,05 |
χ2=0,77 |
p>0,05 |
χ2=28,03 |
p<0,01 |
|
20 |
χ2=0,23 |
p>0,05 |
χ2=0,27 |
p>0,05 |
χ2=28,03 |
p<0,01 |
|
30 |
χ2=4,35 |
p<0,05 |
χ2=5,68 |
p<0,05 |
χ2=26,28 |
p<0,01 |
|
40 |
χ2=20,25 |
p<0,01 |
χ2=18,38 |
p<0,01 |
χ2=25,29 |
p<0,01 |
|
50 |
χ2=28,03 |
p<0,01 |
χ2=28,03 |
p<0,01 |
χ2=23,75 |
p<0,01 |
Также с помощью критерия χ2 мы проверили нулевую гипотезу об отсутствии различий в распределениях количества проникновений в отверстия № 1 и № 2 за 50 проб этапа № 2 (df=49) между испытуемыми выборок №1 и №2 для сопоставления динамики их научения. Результаты для отверстия №1 – χ2= 31,08, p>0,05; результаты для отверстия №2 – χ2= 16,23, p>0,05. Таким образом, нулевая гипотеза принимается в обоих случаях.
Этап № 3. Общая динамика изменения количества проникновений в отверстия № 1 и № 2 у испытуемых трех выборок на этапе № 3 показана на рис. 4 а и 4 б соответственно.
Как видно из графиков, у испытуемых выборок № 1 и № 2 на первых пробах этапа № 3 возрастает количество проникновений и попыток проникновения в оба отверстия по сравнению с количеством проникновений в данные отверстия у этих же выборок на последних пробах этапа № 2. У испытуемых этих двух выборок количество проникновений в отверстие № 1 снижается до 25-й пробы, а далее в среднем составляет 30 (SD=0,83), а количество проникновений в отверстие № 2 снижается до 45-й пробы и на последних пяти пробах в среднем составляет 0 (SD=0,31). У испытуемых выборки № 3 наблюдалась аналогичная динамика, но с противоположными отверстиями: количество проникновений в отверстие № 2 снижается до 25-й пробы, а далее в среднем составляет 30 (SD=0,71), а количество проникновений в отверстие № 1 снижается до 45-й пробы и на последних пяти пробах в среднем составляет 0 (SD=0,44).
Качественная характеристика поведения испытуемых выборки № 1 и № 2.
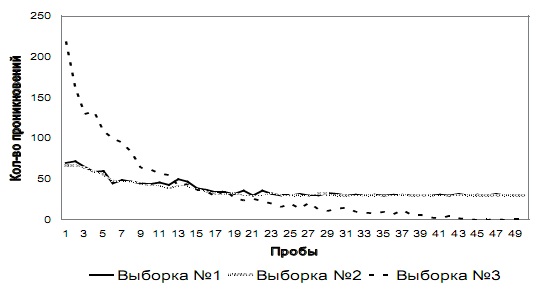
Рис. 4 а. Количество проникновений и попыток проникновения в отверстие № 1 на этапе № 3 у испытуемых трех выборок
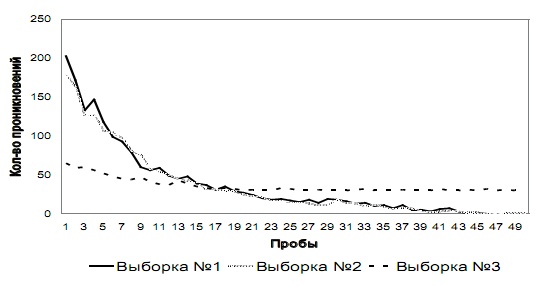
Рис. 4 б. Количество проникновений и попыток проникновения в отверстие № 2 на этапе № 3 у испытуемых трех выборок
На первых пробах этапа № 3 тараканы резко направляются к отверстию № 2 и пытаются пройти через него, упираясь в перегородку отверстия. Далее животные демонстрируют такое же поведение, как и на первых пробах предыдущего этапа. Траектория движения по лабиринту такая же сложная, как на первых сериях этапов № 1 и № 2. В светлой части лабиринта проводят столько же времени, сколько на первых пробах предыдущего этапа. После проникновения в затемненную часть лабиринта ведут себя аналогично предыдущим пробам в этом отсеке. На последних пробах этапа № 3 демонстрируют поведение, аналогичное последним пробам предыдущего этапа, но идут к выходу через другое отверстие. Сразу после попадания в лабиринт направляются к отверстию № 1 и, минуя его, движутся к затемненному отсеку лабиринта, где демонстрируют поведение, аналогичное тому, что проявлялось в данном отсеке на первых пробах. Траектория движения простая – представляет собой прямую линию, ведущую от входа к выходу через отверстие № 1. В светлой части лабиринта тараканы находятся всего несколько секунд.
Поведение испытуемых выборки № 3 на всех пробах этапа № 3 качественно аналогично поведению, продемонстрированному ранее испытуемыми выборки № 1 на соответствующих пробах этапа № 2.
С помощью критерия χ2 с поправкой на непрерывность (df=1) была выполнена проверка нулевой гипотезы об отсутствии различий между эмпирическим распределением количества проникновений в отверстия № 1 и № 2 на каждой пробе этапа № 2 у испытуемых трех выборок и равномерным распределением. В табл. 4 приведены данные по 1, 10, 20, 30, 40 и 50 пробам.
Таблица 4. Значения критерия χ2 при сопоставлении эмпирических распределений количества проникновений в отверстия № 1 и № 2 с равномерным распределением на этапе № 3 у испытуемых трех выборок (достоверные отличия отмечены жирным шрифтом)
|
Пробы |
Выборка №1 (df=1) |
Выборка №2 (df=1) |
Выборка №3 (df=1) |
|||
|
1 |
χ2=63,82 |
p<0,01 |
χ2=50,08 |
p<0,01 |
χ2=58,83 |
p<0,01 |
|
10 |
χ2=1,21 |
p>0,05 |
χ2=1,94 |
p>0,05 |
χ2=3,88 |
p>0,05 |
|
20 |
χ2=1,01 |
p>0,05 |
χ2=0,29 |
p>0,05 |
χ2=0,46 |
p<0,05 |
|
30 |
χ2=3,38 |
p>0,05 |
χ2=2,04 |
p>0,05 |
χ2=5,11 |
p>0,05 |
|
40 |
χ2=20,48 |
p<0,01 |
χ2=22,78 |
p<0,01 |
χ2=20,48 |
p<0,01 |
|
50 |
χ2=28,03 |
p<0,01 |
χ2=28,03 |
p<0,01 |
χ2=28,03 |
p<0,01 |
Как видно из таблицы, для выборок № 1 и № 2 на первой пробе значение критерия выше критического, что обусловлено тем, что испытуемые совершали достоверно больше попыток проникновения в отверстия № 2 (в которое они теперь пройти не могли), нежели проникновений в отверстие № 1. Начиная с 10-й пробы, как показано в таблице, у испытуемых этих двух выборок эмпирическое распределение количества проникновений в отверстия № 1 и № 2 не отличается от равномерного (принимается нулевая гипотеза). Однако, начиная с 40-й пробы и до конца этапа, нулевая гипотеза должна быть отвергнута, так как количество проникновений в отверстие № 1 у испытуемых выборок № 1 и № 2 достоверно преобладает над количеством попыток проникновения в отверстие № 2. Зеркальная ситуация наблюдается у испытуемых выборки № 3: на первой пробе значение критерия выше критического, что обусловлено тем, что испытуемые совершали достоверно больше попыток проникновения в отверстие № 1, начиная с 10-й пробы эмпирическое распределение количества проникновений в отверстия № 1 и № 2 не отличается от равномерного (принимается нулевая гипотеза). Затем, начиная с 40-й пробы и до конца этапа, нулевая гипотеза должна быть отвергнута, так как количество проникновений в отверстие № 2 у испытуемых выборки № 3 достоверно преобладает над количеством попыток проникновения в отверстие № 1.
Также с помощью критерия χ2 мы проверили нулевую гипотезу об отсутствии различий в распределениях количества проникновений в отверстия № 1 и № 2 за 50 проб этапа № 3 (df=49) между испытуемыми выборок № 1 и № 2 для сопоставления динамики их научения. Результаты для отверстия № 1 – χ2= 31,08, p>0,05; результаты для отверстия № 2 – χ2= 16,23, p>0,05. Таким образом, нулевая гипотеза принимается в обоих случаях.
Далее, для определения того, насколько идентично ведут себя испытуемые двух разных выборок в идентичных условиях, с помощью критерия χ2 было сопоставлено распределение количества проникновений в отверстия № 1 и № 2 на этапе № 2 у испытуемых выборки № 1 с распределением количества проникновений в отверстия № 1 и № 2 на этапе № 3 у испытуемых выборки № 3 (df=49). Результаты для отверстия № 1 – χ2= 63,38, p>0,05; результаты для отверстия № 2 – χ2= 49,98, p>0,05. Таким образом, нулевая гипотеза принимается в обоих случаях.
Обсуждение результатов исследования
Из данных, полученных в ходе этапа № 1, можно заключить, что к последним 10 пробам у испытуемых формируется устойчивое научение по прохождению лабиринта через отверстие № 1, как и предполагалось вначале. Это означает, что у насекомых формируется когнитивная карта лабиринта – целостный образ, состоящий из отражения движения собственного тела при локомоции (самоотражения) в отношении пространственных характеристик лабиринта (отражения внешней среды). Следует отметить, что динамика формирования навыка была идентичной у испытуемых всех трех выборок.
В ходе этапа № 2 испытуемые выборки № 1 (с увеличенным размером тела) переучиваются проходить лабиринт через отверстие № 2. На первых пробах животные с увеличенными размерами тела, попадая в уже знакомый лабиринт, начинают осуществлять деятельность, сформированную в научении на предыдущем этапе, – бегут к выходу через отверстие № 1, – и сталкиваются с тем, что данная схема деятельности оказывается нерелевантной условиям, в которых они теперь оказались. Как показывают результаты наблюдения, тараканы в течение нескольких секунд пытаются протиснуться через маленькое отверстие и потом возобновляют эти попытки несколько раз. Вероятно, в результате этих попыток в отражении субъекта количественно накапливаются противоречия между самоотражением – перцептивной схемой собственного тела и динамической разверткой этой схемы при локомоции – и отражением внешней среды (лабиринта) – отражением его представленности в отношении к собственному телу насекомого в виде перцептивного строя (по Барабанщикову, 2002). Затем имеющийся у субъекта целостный образ (когнитивная карта лабиринта) «взрывается» в результате накопления в нем «критической массы» противоречий между самоотражением и внешней средой. Поведенчески это проявляется в возрастании локомоторной активности, передвижении по всей площади лабиринта, т. е. в новом акте ориентировочно-исследовательской деятельности. В результате подобной деятельности тараканы постепенно выстраивают новый перцептивный образ, отличающийся от старого тем, что в нем меняется именно специфика самоотражения субъекта – отражение границ собственного тела – и его отношение к внешней среде.
Однако, как показывают данные, полученные на испытуемых выборки № 2 в ходе этапа № 2, точно таким же образом формируется научение прохождения лабиринта через отверстие № 2 и в том случае, если не увеличиваются размеры тела, а уменьшаются размеры отверстия № 1. Это отражается и в отсутствии достоверных отличий в проникновении в оба отверстия между испытуемыми выборок № 1 и № 2, и в наблюдаемом поведении, которое также идентично у насекомых этих двух выборок.
Данный факт можно объяснить следующим образом. По-видимому, тараканы отражали лишь собственную «неспособность» проникать в отверстие № 1 и следовать прежним путем к выходу – для них «не важно», чем обусловлена эта неспособность: увеличением границ их собственного тела или уменьшением отверстия. При формировании навыка у этих насекомых изменился весь целостный образ ситуации, а не какая-либо его часть (самоотражение или отражение внешней среды) в отдельности. Соответственно, эти изменения затрагивают и специфику самоотражения насекомых как неотделимую часть этого образа. Тем не менее, объективно изменения пространственных характеристик размеров тела учитывались испытуемыми выборки № 1 при ориентации в пространстве, – в ином случае насекомые не смогли бы достичь выхода. Новая перцептивная схема собственного тела сложилась у этих животных именно в процессе тактильного контакта с объектами внешнего мира (отверстиями в лабиринте), т. е. представлена в форме непосредственных переживаний, возникающих в процессе физического взаимодействия с этими объектами. Пенопластовый куб, прикрепленный к поверхности их тела, играл сразу две роли: роль зонда, очерчивающего границы тела субъекта и поставляющего ему кинестетические ощущения об этих границах, и роль фактора, определяющего свободу передвижения субъекта. У испытуемых выборки № 2 ситуация была аналогичной за тем исключением, что роль зонда выполняло их собственное тело, которое оставалось прежним.
Данные этапа № 3, полученные на испытуемых выборок № 1 (увеличенное тело) и № 2 (исходное тело), свидетельствуют, что новое научение в случае повторного изменения условий среды у тараканов происходит так же, как и в первый раз: идентична и динамика научения по количеству проникновений и попыток проникновения в отверстия, и наблюдаемое поведение. Это говорит о том, что эти насекомые не способны переносить уже сформированный навык из одной ситуации в другую, т. е. когнитивные карты, выстраиваемые ими, весьма узки, что согласуется с данными других авторов (Филиппова, 2004).
Данные, полученные на испытуемых выборки № 3, показывают, что поведение насекомых никак не меняется даже при увеличении границ их тела в том случае, если это не препятствует прохождению лабиринта уже выученным путем. По-видимому, в таком случае не меняется и когнитивная карта – целостный образ ситуации, а также его составные части (самоотражение и отражение внешней среды). Однако на этапе № 3, когда отверстие № 1 в лабиринте было уменьшено до непроходимости для испытуемых выборки № 3 с измененной формой тела, они начали переучиваться, причем динамика научения и поведение идентичны тем, что демонстрировали испытуемые выборки № 1 в ходе этапа № 2 в идентичных условиях, также с увеличенным размером тела. Эти факты еще раз подтверждают уже сформулированный тезис о том, что целостный образ ситуации, а также самоотражение как его составной компонент, у этих животных формируется и меняется лишь в процессе деятельности и взаимодействия (в данном случае тактильного) с объектами внешней среды.
Заключение
В результате экспериментального исследования установлено, что у тараканов как представителей низшего уровня перцептивной стадии развития психики отражение внешней среды и самоотражение представлены как слитные в структуре единого образа. На данной стадии животные уже способны отражать пространственные характеристики своего тела (границы своего тела) в виде чувственных переживаний, возникающих в процессе взаимодействия с объектами внешнего мира. Кроме того, эти животные при изменении размеров их тела (увеличении) могут путем научения через взаимодействие с объектами внешней среды отражать эти изменения и учитывать их при ориентации во внешнем пространстве.
Таким образом, гипотеза настоящего исследования была подтверждена.
На низшем уровне перцептивной стадии развития психики самоотражение и отражение внешней среды только начинают разделяться в структуре единого перцептивного образа, и изменения, затрагивающие какую-либо одну часть данного образа, неизбежно касаются и других его частей. Субъект еще не способен отражать свое тело как целое, но лишь его отдельные пространственные характеристики. Однако это самоотражение уже поддается изменению путем научения, осуществляющемуся лишь в контакте с внешней средой в процессе деятельности.
Перспективным представляется проведение аналогичных сравнительно-психологических экспериментальных исследований на животных, относящихся как к более высоким, так и более низким стадиям развития психики.