Элементарное мышление животных — это способность оперировать эмпирическими законами, связывающими предметы и явления внешнего мира, оперировать этими законами в новой для животного ситуации (на основании врожденных инстинктов или ранее сформированных навыков) для построения программы адаптивного поведенческого акта (Крушинский, 2009). Соответственно, именно благодаря мышлению животные осуществляют индивидуальную гибкую адаптацию к тем уникальным условиям среды, в которых они оказались в данный момент времени, с учетом информации о себе и об окружающей среде.
Обобщая взгляды различных авторов на природу элементарного мышления животных, З.А. Зорина и И.И. Полетаева выделили основные аспекты проявления данного феномена (Зорина, Полетаева, 2003). К их числу относится способность осуществлять обобщение и абстрагирование, т. е. выделять и фиксировать относительно устойчивые, инвариантные свойства предметов и их отношений в процессе обучения и приобретения опыта.
В своей периодизации эволюции психики А.Н. Леонтьев относил животных, обладающих мышлением, к отдельной стадии — стадии интеллектуальной психики, предшествующей возникновению человеческого сознания (Леонтьев, 1972). Согласно классическим теориям научения, мышление является достоянием психики только теплокровных позвоночных — млекопитающих и птиц (Филиппова, 2012).
Вместе с тем существует традиция рассмотрения мышления как механизма интеграции различных когнитивных компонентов (элементов интеллекта) (Веккер, 1998). В этом смысле мышление является неотъемлемым атрибутом любого когнитивного процесса — даже у представителей более ранних стадий эволюции психики. С точки зрения данного подхода, предпосылкой формирования мышления является способность животного переносить ранее приобретенный навык в новую ситуацию (Филиппова, 2012).
Традиционно рептилии считаются ограниченными с точки зрения своих когнитивных способностей в сравнении с млекопитающими и птицами (Burghardt, 1977). Однако современные экспериментальные данные свидетельствуют о наличии у них когнитивных способностей, аналогичных тем, что обнаруживаются у теплокровных позвоночных (Wilkinson A., Huber, 2012). В частности, было показано, что ящерицы способны переносить ранее приобретенный опыт в новую ситуацию при решении дифференцировочной задачи (там же), а также способны формировать навык путем имитации и подражания (Noble et al., 2014).Рептилии являются ключевым классом в развитии понимания эволюции когнитивных способностей среди амниотов. Изучение сходств и различий в их познавательных процессах может информировать о гомологиях и аналогах когнитивных механизмов у амниотов в целом (Matsubara et al., 2017). В частности, это касается и феномена схемы тела — процесса ее формирования и модификации у рептилий.
Между тем, до сих пор не было получено достоверных данных, свидетельствующих о способности рептилий переносить ранее полученный опыт в новые ситуации. С другой стороны, крысы способны к обобщению приобретенного опыта и переносу его в новые ситуации — формированию правил (Murphy et al., 2008).
Целью настоящего исследования является сравнительно-психологический анализ особенностей решения задачи на перенос ранее приобретенного навыка в новые условия у рептилий на примере синеязыких сцинков (Tiliquagigas) и у млекопитающих на примере серых крыс (Rattusnorvegicus).
Гипотеза исследования: хотя оба вида животных демонстрируют способность к переносу ранее приобретенного навыка в новые ситуации, этот феномен качественно отличается у этих видов.
В конкретном смысле мы полагаем, что сцинк способен научаться лишь учету увеличенных границ собственного тела, в то время как крыса может гибко модифицировать свое поведение в зависимости от ситуативного соотношения границ ее тела и границ внешних объектов.
Необходимо отметить, что часть данных, обсуждаемых в настоящей статье, ранее была представлена в других работах (Хватов и др., 2016 a; 2016 b), однако здесь мы также приводим и анализируем новые эмпирические данные (полученные в 5-й серии эксперимента — см. ниже).
Методика исследования
Испытуемые животные.
Сцинки. 8 сцинков: 6 самцов и 2 самки (половых различий в поведении в рамках эксперимента выявлено не было) гигантских синеязыких сцинков Tiliquagigas (далее — сцинк), взрослые, наивные особи, пойманные в дикой природе и наблюдавшиеся в лабораторных условиях около 1 года.
Крысы. В эксперименте было использовано 8 крыс породы Long-Evans, самцы в возрасте от 2 до 6 мес.
Общая характеристика экспериментальной установки. Для обоих видов был использован сходный тип экспериментальных установок (небольшие отличия описаны ниже), представлявших собой квадратные террариумы. Внутри террариума, в центре, располагался проблемный ящик квадратной формы. В каждой из боковых стенок ящика было по одному отверстию. Внутри проблемного ящика находился корм для животного. Испытуемому необходимо было проникнуть головой через одно из отверстий внутрь проблемного ящика для достижения корма.
Независимыми переменными являлись:
• границы тела животного;
• границы объектов внешней среды — диаметр отверстий в экспериментальной установке.
Границы тела животных увеличивались путем крепления на их тело различных инородных объектов: с помощью приклеивания (сцинки) (рис. 1), с помощью монтирования в череп (крысы) (рис. 1).

В эксперименте использовалось три типа диаметра отверстий:
• малые отверстия (далее — отверстия S) — через них животное не могло достичь приманку или попасть в укрытие;
• средние отверстия (далее — отверстия M) — через них лишь животное с естественными, но не увеличенными границами тела могло достичь приманку или попасть в укрытие;
• большие отверстия (далее — отверстия L) — через них животное и с естественными, и с увеличенными границами тела могло достичь приманку или попасть в укрытие.
Зависимой переменной являлось поведение животного в ходе взаимодействия с объектами внешней среды для решения различного рода экспериментальных задач — количество проникновений в различные типы отверстий. При этом за одну попытку проникновения считалась такая ситуация, когда животное упиралось в границы отверстия объектом, закрепленным у него на голове, совершая возвратно-поступательные движения мордой и/ или роющие (буксующие) движения передними лапами.
С каждой экспериментальной группой проводилось несколько экспериментальных серий, в каждой их которых изменялись параметры независимых переменных. Каждая серия состояла из 20 проб.
Экспериментальная установка для сцинков была организована в прямоугольном вертикальном террариуме (размер террариума: 450x450x620 мм). Внутри террариума располагался проблемный ящик (размер ящика: 150x150x220 мм) (рис. 2). Также в террариуме располагалась поилка с водой (рис 2).

Кормление животного осуществлялось только внутри экспериментальной установки таким образом, что для достижения корма животному необходимо было проникать головой в одно из отверстий. Кормушка (пластиковая подставка диаметром 100 мм на ножках высотой 10 мм) с приманкой помещалась в центр экспериментальной установки в начале каждой пробы.
Экспериментальная установка для крыс представляет собой стеклянный террариум квадратной формы (со стороной 720 мм) с пятью отсеками: центральным отсеком квадратной формы (со стороной 200 мм) и четырьмя отсеками в форме равнобедренных трапеций, обрамляющими его. Обрамляющие отсеки сообщались между собой аркообразными отверстиями в форме усеченного круга (диаметром 100 мм), расположенными в центре боковых сторон отсеков (рис. 3). Центральный отсек сообщался с каждым из четырех обрамляющих отсеков круглыми отверстиями (диаметром 80 мм), располагавшимися по центру каждой из сторон отсека на высоте 5 мм от пола.
Ниже приведем описание экспериментальных серий. Каждая экспериментальная серия состояла из 20 проб. В начале экспериментальной пробы животное располагалось напротив отверстия № 1.
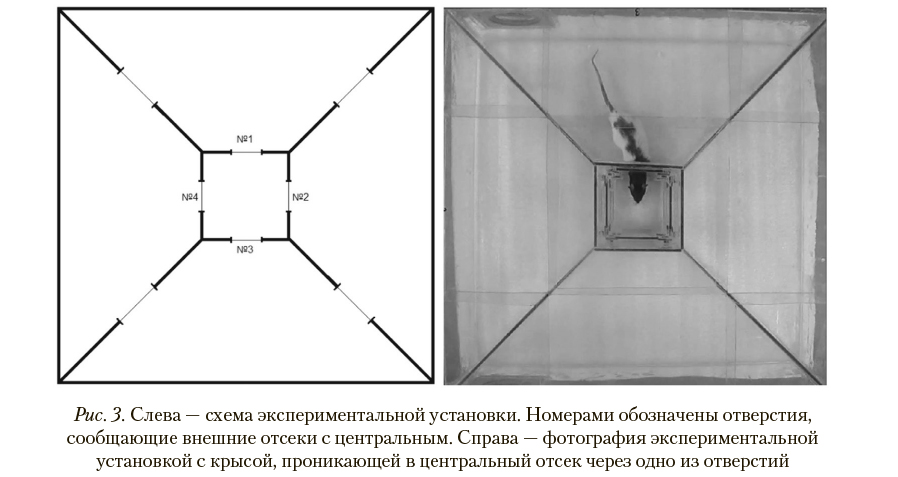
Таблица 1
Схема экспериментального плана (пояснения см. в тексте)
|
Экспериментальные серии |
Независимая переменная 1 — границы тела животных |
Независимая переменная 2 — схема расположения отверстий в экспериментальной установке |
|
Серия 1 |
Естественные |
№ 1 — M; № 2 — S; № 3 — S; № 4 — S |
|
Серия 2 |
Увеличенные |
№ 1 — M; № 2 — S; № 3 — L; № 4 — S |
|
Серия 3 |
Увеличенные |
№ 1 — M; № 2 — L; № 3 — M; № 4 — S |
|
Серия 4 |
Увеличенные |
№ 1 — M; № 2 — M; № 3 — M; № 4 — L |
|
Серия 5 |
Рандомные в каждой пробе |
Рандомная в каждой пробе |
Серия 1. Схема расположения отверстий: № 1 — M; № 2 — S; № 3 — S; № 4 — S. Таким образом, животное могло решить задачу путем проникновения лишь в одно конкретное отверстие. Задачей серии являлось формирования у животного навыка быстрейшего проникновения во второй отсек или внутрь проблемного ящика для достижения приманки. Экспериментальная серия продолжалась до тех пор, пока животное в течение пяти проб подряд не совершало неуспешных попыток проникновения в отверстие S и сразу же проникало во второй отсек через отверстие M.
Серия 2. Схема расположения отверстий № 1 — M; № 2 — S; № 3 — L; № 4 — S. Границы тела животных увеличивались. Соответственно, животное не могло проникнуть в отсек 2 ранее выученным маршрутом не из-за изменения характеристик внешней среды, а из-за изменения характеристик его тела. Критерии сформированности навыка были прежними.
Серия 3. Схема расположения отверстий: № 1 — M; № 2 — L; № 3 — M; № 4 — S. Границы тела животного оставались увеличенными, а схема расположения отверстий снова менялась: проницаемое отверстие L теперь перемещалась в новое место (отверстие, ранее имевшее диаметр L, теперь уменьшалось до M). Животному было необходимо реорганизовывать свой прежний навык. Критерии сформированности навыка были прежними.
Серия 4. Схема расположения отверстий: № 1 — M; № 2 — M; № 3 — M; № 4 — L. Границы тела животного были увеличены, а схема расположения отверстий менялась. В данной серии проверялось, способно ли животное перенести ранее сформированный навык в новые условия. Критерии сформированности навыка были прежними.
Мы считали, что у животного сформировался навык учета измененных границ своего тела в том случае, если в третьей серии оно совершало достоверно меньше попыток проникновения в средние отверстия, нежели в серии 2. Такого рода выученное поведение может указывать на наличие у животного способности переносить ранее приобретенный опыт «непроницаемости» отверстия в новую измененную ситуацию.
Серия 5. В данной серии в каждой пробе осуществлялось изменение случайным образом границ тела животных — они либо оставались естественными, либо были увеличены за счет специальных приспособлений. Расположение отверстий в экспериментальной установке также варьировалось случайным образом в каждой пробе (между отверстиями типов M и L), при условии, что хотя бы одно отверстие являлось M, а одно — L. Задачей серии являлось установить, насколько гибко животные способны варьировать свое поведение в зависимости от условий окружающей среды и особенностей границ собственного тела, т. е. способны ли животные решать новую задачу на основе ранее усвоенных закономерностей.
Результаты эксперимента
Серия 1. Крысы с естественными границами тела в ходе всей экспериментальной серии не совершали неуспешных попыток проникновения в отверстия типа S, но достигали приманку только через отверстия M. Сцинки в ходе экспериментальной серии начинали совершать все меньше неуспешных попыток в отверстия S (№ 2, 3 и 4) и все чаще напрямую достигать приманку через отверстие M — № 1. У сцинков на первых 5 пробах серии доля попыток проникновения в отверстие № 1 составляла 17%, на последних 5 пробах — 100%; Х2=116,15; df=3; p<0,01.
Серия 2. И крысы, и сцинки к концу экспериментальной серии стали чаще совершать попытки проникновения в отверстия № 3 и реже — в отверстия № 1, 2 и 4. Крысы за первые 5 проб совершили 48 неуспешных попыток проникновения в отверстия M, за последние 5 проб — 0; х2=34,91; df=3; p<0,01. Сцинки за первые 5 проб совершили 108 неуспешных попыток проникновения в отверстия M, за последние 5 проб — 0; х2=68,59; df=1; p<0,01. Результаты экспериментальной серии 2 представлены в табл. 2.
Таблица 2
Общее количество попыток проникновения, совершенных всеми животными выборки,
за первые и последние 5 проб серии
|
Пробы |
Попытки проникновения в отверстия за приманкой у крыс |
Попытки проникновения в отверстия за приманкой у сцинков |
||
|
В отверстия M |
В отверстия L |
В отверстия M |
В отверстия L |
|
|
Пробы 1—5 |
48 |
40 |
108 |
40 |
|
Пробы 16—20 |
0 |
40 |
0 |
40 |
Серия 3. Начиная с данной серии, крысы не совершали более неуспешных попыток проникновения в отверстия M. Сцинки же за первые 5 проб совершили 53 неуспешные попытки проникновения в отверстия M, за последние 5 проб — 0; х2=37,90; df=1; p<0,01. При этом за первые 5 проб данной серии сцинки совершили значительно меньше неуспешных попыток проникновения в отверстия M, нежели за первые 5 проб серии 2; х2=6,58; df=1; p<0,05. Однако за первые 5 проб данной серии сцинки совершали больше неуспешных попыток проникновения в отверстия M, нежели крысы за первые 5 проб данной серии Х2=37,90; df=1; p<0,01. Результаты экспериментальной серии 3 представлены в табл. 3.
Таблица 3
Общее количество попыток проникновения, совершенных всеми животными выборки,
за первые и последние 5 проб серии
|
Пробы |
Попытки проникновения в отверстия за приманкой у крыс |
Попытки проникновения в отверстия за приманкой у сцинков |
||
|
В отверстия M |
В отверстия L |
В отверстия M |
В отверстия L |
|
|
Пробы 1—5 |
0 |
40 |
53 |
40 |
|
Пробы 16—20 |
0 |
40 |
0 |
40 |
Серия 4. Как и в предыдущей серии крысы не совершали более неуспешных попыток проникновения в отверстия M. Сцинки же за первые 5 проб совершили 15 неуспешных попыток проникновения в отверстия M, за последние 5 проб — 0; х2=12,95; df=1; p<0,01. При этом за первые 5 проб данной серии сцинки совершили значительно меньше неуспешных попыток проникновения в отверстия M, нежели за первые 5 проб серии 3; х2=12,29; df=1; p<0,01. Однако за первые 5 проб данной серии сцинки, как и в предыдущей серии, совершали больше неуспешных попыток проникновения в отверстия M, нежели крысы за первые 5 проб данной серии х2=37,90; df=1; p<0,01. Результаты экспериментальной серии 4 представлены в табл. 4.
Таблица 4
Общее количество проникновений, совершенных всеми животными выборки,
за первые и последние 5 проб серии
|
Пробы |
Попытки проникновения в отверстия за приманкой у крыс |
Попытки проникновения в отверстия за приманкой у сцинков |
||
|
В отверстия M |
В отверстия L |
В отверстия M |
В отверстия L |
|
|
Пробы 1—5 |
0 |
40 |
15 |
40 |
|
Пробы 16—20 |
0 |
40 |
0 |
40 |
Серия 5. Теперь рассмотрим данные эксперимента, в котором осуществлялась рандомизация границ тела животных и схем расположения отверстий в экспериментальной установке. Результаты экспериментальной серии 5 представлены в табл. 5.
Таблица 5
Общее количество попыток проникновения в различные типы отверстий за все 160
(общее количество проб, совершенных всеми животными конкретной группы в серии 5)
|
Проникновения/по- пытки проникновений |
Границы тел крыс |
Границы тел сцинков |
||
|
Естественные |
Увеличенные |
Естественные |
Увеличенные |
|
|
В отверстия M |
33 |
0 |
3 |
2 |
|
В отверстия L |
39 |
88 |
84 |
73 |
Из 160 экспериментальных проб, проведенных с крысами (общее количество проб, совершенных всеми животными конкретной группы в серии 5), в 72 случаях границы тела крысы оставались естественные, в 88 — увеличивались за счет специальных приспособлений. Эмпирическое распределение количества проникновений и попыток проникновения в различные типы отверстий при естественных границах тел достоверно отличается от аналогичного распределения при увеличенных границах тел (х2=50,81; df=1; p<0,01).
У сцинков в 160 экспериментальных пробах в 87 случаях границы тела были естественные, в 73 — увеличенные. Эмпирическое распределение количества проникновений и попыток проникновения в различные типы отверстий при естественных границах тел не имеет достоверных отличий от аналогичного распределения при увеличенных границах тел (х2=0,08; df=1; p>0,05). При этом у сцинков не было выявлено динамики в выборе типа отверстий: в первых 5 пробах сцинки совершили 0 попыток проникновения в отверстия M и 40 попыток проникновения в отверстия L, на последних 5 пробах — одну попытку проникновения в отверстия M и 40 попыток проникновения в отверстия L (табл. 6) — распределения не имеют достоверных отличий (х2=0,99; df=1; p>0,05).
Таблица 6
Результаты экспериментальной серии 5 у сцинков. Общее количество проникновений, совершенных всеми животными выборки за первые и последние 5 проб серии
|
Пробы |
Попытки проникновения в отверстия за приманкой у сцинков |
|
|
В отверстия M |
В отверстия L |
|
|
Пробы 1—5 |
0 |
40 |
|
Пробы 16—20 |
1 |
40 |
Эмпирическое распределение количества проникновений в отверстия M и L у крыс при естественных границах тел в серии 5 достоверно отличается от аналогичного распределения у сцинков в серии 5 (х2=40.41; df=1; p<0.01).
Обсуждение результатов
Полученные данные свидетельствуют о том, что крысам не требовалось формировать навык учета естественных границ их тел (вероятно, потому, что такой навык был сформирован ранее), так как в ходе первой экспериментальной серии крысы не демонстрировали неуспешных попыток проникновения в отверстия типа S. У сцинков же количество неуспешных попыток проникновения в отверстия S снижалось от начала к концу серии. Соответственно, у рептилий навык учета естественных границ своего тела складывался в ходе эксперимента.
При увеличении границ тела в начале второй серии и крысы, и сцинки продемонстрировали множественные попытки проникновения в отверстия M (теперь непроницаемые для них). К концу серии таких попыток проникновения животные больше не осуществляли.
У сцинков на первых 5-ти пробах серий 3 и 4 наблюдалось возрастание количества попыток проникновения в отверстия M в сравнении с последними 5 пробами предыдущей серии соответственно. Однако в начале каждой следующей серии таких попыток становилось все меньше.
Результаты третьей и четвертой серии продемонстрировали существенную разницу между результатами крыс и сцинков. Несмотря на то, что оба вида животных после увеличения размеров их тела оказались способны обучиться использовать именно большое
отверстие (во второй серии), только крысы, но не сцинки, без дополнительного обучения смогли перенести этот навык в новые экспериментальные ситуации (в третьей и четвертой сериях), в которых меняли местоположение этого отверстия. Сцинкам для выбора большого отверстия при изменении его местоположения требовалось дополнительное обучение.
Ключевое значение имеют результаты 5-й серии. Крысы продемонстрировали способность гибко менять свое поведение в зависимости как от условий внешней среды, так и от особенностей границ собственных тел. Это свидетельствует о том, что крысы не просто научились воспринимать увеличенные границы собственного тела, но и усвоили эмпирическую закономерность соотношения границ своего тела и отверстий в экспериментальной установке.
Сцинки же в 5-й серии, как при увеличенных границах тела, так и при естественных, предпочитали достигать приманку через отверстие L. Это означает, что у сцинков сформировался лишь навык учета увеличенных границ своего тела инвариантно по отношению к тому увеличены ли объективно границы их тел или не увеличены. В отличие от крыс сцинки не усвоили вышеописанной эмпирической закономерности, в силу чего их поведение оказалось ригидным.
Таким образом, можно констатировать, что именно у крыс в отличие от сцинков наблюдается процесс мышления при решении данной экспериментальной задачи, проявлением которого стало формирование обобщенного психического образа (схемы собственного тела в отношении к характеристикам внешних объектов) — в данном случае модификации восприятия границ собственного тела, — а также способность обнаруживать общие свойства в окружающих предметах и общие закономерности в различных ситуациях (Фирсов, 1993; Зорина, Полетаева, 2003).
Тот факт, что сцинки переносят ранее приобретенный опыт в новые ситуации — хотя и не с такой эффективностью и скоростью, как крысы, — свидетельствует в пользу наличия у них предпосылок формирования мышления.
Мы можем предположить, что именно в ходе эволюции ранних высших позвоночных (амниот), с которыми наибольшее сходство имеют современные пресмыкающиеся, закладывались предпосылки формирования как морфофизилогической, так и психической организации более поздних видов. Выдвинутое предположение подтверждается также данными о том, что аналогичная способность к переносу ранее приобретенного навыка обнаруживается у других представителей пресмыкающихся — змей, но отсутствует у более древних представителей позвоночных — земноводных (Хватов, Харитонов, 2017).
Выводы
Синеязыкие сцинки (Tiliquagigas) и серые крысы (Rattusnorvegicus), как представители рептилий и млекопитающих соответственно, демонстрируют способность к переносу ранее приобретенного навыка. Однако сцинки формируют правило учета лишь увеличенных границ тела, крысы же способны гибко соотносить границы своего тела с характеристиками внешних объектов и формируют более сложное правило выбора различных типов отверстий в зависимости от того, увеличены границы их тела или нет в каждой конкретной ситуации.
Финансирование
Работа поддержана грантом РФФИ (проект № 17-06-00832-а).