Поведение живых организмов направлено на получение результатов – нужных соотношений со средой, и история этих соотношений со средой, фиксирована в структуре индивидуального опыта в виде элементов опыта разного возраста (Александров, 1995, 2006; Швырков, 1987, 2006; Шевченко и др., 1997). Акустические параметры среды (звуки) определяются как экологической нишей вида, так и особенностями среды, в которой живет конкретный индивид. Связанные с результатами поведения акустические параметры среды (акустические события) оказываются, таким образом, биологически значимыми, и через них возможна актуализация элементов опыта. Сравнительный анализ палеогенеза акустических структур (Харитонов, 2004) позволяет предположить, что некоторые звуки, становясь акустическими событиями, являются одинаково значимыми для представителей разных видов (например, для людей и для крыс), тогда как другие акустические события являются важными только для представителей одного вида, а какие-то – только для группы или конкретного индивида. Это определяется сходством и различиями в наборах элементов индивидуального опыта у индивидов разных видов и индивидуальными различиями у особей в пределах вида. Следовательно, акустические параметры среды (звуки) и акустические события (значимые звуки) могут быть использованы для тестирования наличия тех или иных элементов опыта у индивида и для определения отношений между элементами опыта.
Мы исходили из предположения, что разные звуки будут актуализировать разные элементы опыта, что приведет к изменениям в поведении и отразится в амплитудно-временных параметрах электрической активности мозга (Гаврилов, 1987; Максимова, Александров, 1987).
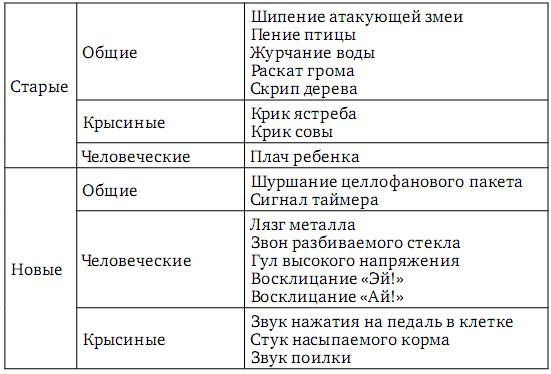
Поскольку элементы опыта имеют разный возраст и разную акустическую составляющую, поэтому появление разных звуков должно приводить к актуализации элементов опыта разного возраста. С этой целью среди звуков были выбраны те, которые могли бы актуализировать как сходные (новые и старые) элементы опыта у человека и у крыс, так и разные (новые и старые) – у человека и у крыс. Мы выбрали 18 звуков, которые, исходя из нашего предположения, разделили на следующие 6 групп:
Все звуковые файлы были отформатированы так, чтобы их длительность не превышала 1 с (поскольку длительность побежки крысы от педали к кормушке в среднем была около 1 с), но в то же время звук оставался узнаваем.
В инструментальном пищедобывательном поведении крысы должны были нажимать на педаль для получения порции пищи в кормушке. При побежке от педали к кормушке появлялся тот или иной звук. Предъявления звуков чередовались с «пустыми» пробами без звука. Регистрировали суммарную электрическую активность мозга над моторной, зрительной, слуховой и лимбической областями коры, для чего использовали хлорсеребряные электроды, вживленные эпидурально с референтом над носовыми пазухами.
Были проведены психофизиологические эксперименты на 4 крысах (самцах Wistar), предварительно обученных добывать себе пищу нажатием на педаль. Для этого и подобных экспериментов было разработано программное обеспечение, которое позволило:
- автоматизировать подачу звуков по определенному сценарию в нужные моменты времени при реализации индивидом определенного поведенческого акта;
- синхронизировать регистрируемые данные (отметки поведения и подачи звука с суммарной электрической активностью мозга).
Аппаратно-программный комплекс включает следующие элементы:
- Экспериментальная клетка в виде куба со стороной 60см, в углах которой рас положены педаль и кормушка. Для определения пересечения крысой середины стенки – начала предъявления звука – были установлены фотоэлектрическая пластина ФЭП и излучающий светодиод (рисунок 1).
- Два АЦП для сбора сданных: National Instruments USB-6008 (NI) для автоматизации подачи звуков и Lcard DL-120 для сбора поведенческих и ЭЭГ данных.
- Программы: сбора данных DigiScope (Ds), подачи звука (Е. Нармухамедова), обработки ЭЭГ (Psy 2.2) (А. Анашкин).
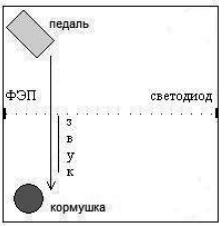
Рис. 1. Схема экспериментальной клетки (вид сверху) Стрелкой показан акт побежки от педали к кормушке, во время которого появлялся звук, как только крыса пересекала середину стенки, что фиксировалось фотоэлектрической пластиной. Рядом линией условно показана длительность звука пропорционально времени реализации поведенческого акта.
Функционирование комплекса происходит по следующей схеме: сигналы с датчиков о нажатии на педаль, опускании головы в кормушку и пересечении середины стенки поступают на АЦП (NI), где они преобразуются в цифровую форму. Программа на компьютере считывает сигналы и при наступлении определенных условий воспроизводит звук. В момент запуска звука обратно на АЦП подается соответствующая каждому звуку метка. Сигнал с отметками, вместе с сигналом от датчиков нажатия на педаль или опускания головы в кормушку и энцефалографическими данными поступает на второй АЦП (Lcard) и после преобразования – в регистрирующую программу.
Для подачи звука в среде программирования NI LabView нами была разработана специальная программа, которая позволяет выбирать звуковые файлы, которые следует воспроизводить. На вход программы поступают данные с АЦП о текущем поведении: нажатии на педаль, пересечении середины стенки, опускании головы в кормушку. При условии, что крыса нажала на педаль и пересекла середину стенки, программа случайным образом выбирает один из звуков и проигрывает его. В этот же момент с помощью ЦАП-АЦП подается отметка в программу сбора данных о проигрываемом звуке.
Проведено несколько экспериментальных серий, в которых: 1) звуки появлялись в инструментальном пищедобывательном поведение:
- при побежке крысы к кормушке на середине пути между педалью и кормушкой;
- при нажатии крысы на педаль (для исключения вероятности того, что некоторые звуки продолжают звучать на протяжении всего этапа анализируемого поведения – побежки от середины стенки к кормушке); 2) звуки предъявлялись при фиксации крысы:
- в гамаке (для исследования вызванных потенциалов при ограничении движений);
- под наркозом (без поведения). Для каждой из серий программа для подачи звука была модифицирована в соответствии с задачами.
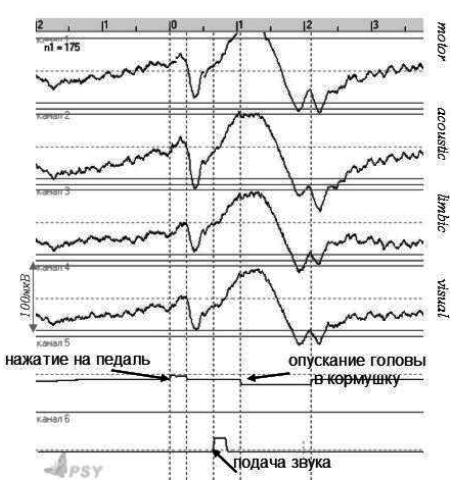
Рис. 2. Усредненные ЭЭГ потенциалы от отметок поведения – вертикальные пунктирные линии (Rat#4). N = 175 – число реализаций в усреднении; наверху – шкала времени в секундах
Полученные данные обрабатывались в программе Psy 2.2 (А. Анашкин), которая позволяет отмечать границы поведенческих актов и усреднять ЭЭГ от этих отметок. Для разделения отметок о звуках на разные категории в зависимости от самого звука была написана программа в системе LabView, которая обрабатывала файлы с отметками и из одного файла со всеми отметками создавала отдельный файл для каждого звука. Используя полученные отметки, в программе Psy 2.2 усреднялись суммарные потенциалы мозга от отметок нажатия на педаль, опускания головы в кормушку и от момента подачи звука.
Получены следующие предварительные результаты: реализациям и сменам актов поведения соответствуют негативные и позитивные ЭЭГ-колебания сходные по конфигурации и амплитуде во всех отведениях (рисунок 2). Появление разных звуков при реализации одного и того же поведенческого акта «разбило» этот акт на два субакта, что отразилось в появлении дополнительного ЭЭГ-компонента – «ступеньки», который возник в результате появления дополнительного позитивного колебания во время негативизации при побежке крысы к кормушке (отмечен звездочкой на рисунке 2). Этот компонент сохранялся и в реализациях без предъявления звука, что можно объяснить тем, что он связан с ожиданием звука.
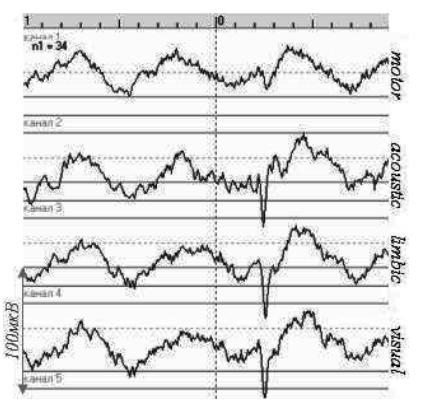
Рис. 3. Усреднение от момента предъявления звука «Звон меча» – вертикальная пунктирная линия – у крысы в гамаке (Rat#1). N = 34 – число реализаций в усреднении; наверху – шкала времени: одно деление – 100 мс
Значимых различий в ЭЭГ в момент предъявления разных звуков выявлено не было. Только на один звук («звон меча») при регистрации ЭЭГ у крысы, фиксированной в гамаке, был выявлен выраженный во всех отведениях потенциал (отмечен на рисунке 3). Для оценки изменений в поведении рассматривались временные интервалы от окончания нажатия на педаль до опускания головы в кормушку. Для анализа скорости реализации поведенческих актов была разработана специальная програм ма. Статистическая обработка данных проводилась в пакете Statistica с помощью однофакторного дисперсионного анализа. Были определены звуки, при появлении которых время реализации акта побежки крысы от педали к кормушке значимо отличалось от других. Оказалось, однако, что для разных крыс такие звуки были разные. При этом их появление могло по-разному сказываться на скорости реализации исследуемого поведенческого акта: у одних крыс замедлять, у других – ускорять.
Анализ времени реализации актов поведения в зависимости от групп звуков выявил некоторые значимые отличия. Выделилась группа «Человеческие новые» звуки, которая значимо отличается от групп «Крысиных новых», «Человеческих старых» звуков и «Без звука». При появлении звуков из группы «Человеческие новые» сокращалось время реализации анализируемого акта поведения, во время которого они появлялись. Также установлено, что среднее время реализации акта побежки к кормушке, когда звук не появлялся, было значимо больше времени реализации того же акта при появлении звуков.
Полученные предварительные данные можно, по-видимому, объяснить различиями в индивидуальном опыте крыс, актуализацией разных элементов опыта в зависимости от значения звука. У разных животных появление одних и тех же звуков по-разному сказывалось на временных (скоростных) характеристиках реализации поведенческих актов. Однако в настоящее время недостаточно данных для выводов о закономерностях в отношениях между элементами индивидуального опыта, возникающими при актуализации элементов опыта разными акустическими событиями (значимыми звуками).
В последующих экспериментах предполагается увеличить не только объем выборки животных, но и репертуар звуков, проверить «качество» звуков, искусственно придать значение некоторым звукам, ранее незначимым для животного. Разработанный АПК может быть легко перенастроен и использован для выполнении разнообразных аналогичных психофизиологических исследований. Данная работа выполнена в рамках более широкого сравнительного исследования восприятия звуков человеком и животными.