Информация об окружающем мире представлена в памяти не только репрезентациями в виде отдельных объектов, ландшафтов, ситуаций, действий, событий, но и в форме их обобщений, категорий. Категоризация – жизненно важный когнитивный процесс, позволяющий правильно идентифицировать любой объект, событие и связывать его с соответствующим поведением. Запоминание новых объектов (стимулов) и их опознание также предполагает участие процесса категоризации, направленного как на сами объекты, так и на действия, которыми требуется на них отвечать. Изучение нейрональных механизмов категоризации ведется в трех основных направлениях: изучаются процессы и структуры мозга, вовлеченные в репрезентации категорий самих объектов; исследуется, как выбирается действие или поведение, которое связывается с объектом, и как оно обобщается; изучается, как влияет тренировка, память на динамику процесса формирования категории. Исследования проводятся на животных, часто с регистрацией нейронной активности, и на людях в основном с применением метода функциональной магнитно-резонансной томографии (фМРТ).
Категоризация – критический и динамический аспекты принятия решения, механизм которого еще не ясен. Одна из тем, которая интересует исследователей, – какой вклад в процессы категоризации вносят различные области коры мозга. По этой теме мнения разделились. Крайнюю позицию занимают исследователи, утверждающие, что формирование категорий связано с функцией фронтальной коры, а височная кора включена в обработку только физических параметров стимула (Freedman et al., 2003). Промежуточная позиция выразилась в заключении, что фронтальная кора включена в анализ и формирование абстрактных категорий (Grinband et al., 2006). Авторы, используя метод фМРТ и варьируя неопределенностью категорий, выявили участие двух областей фронтальной доли в процессах категоризации. Одна обнаружена в медиальной фронтальной коре (ВА8), другая в передней фронтальной коре – передней инсулярной коре (anterior insula – АI). Они показали связь активности в ВА8 и АI с изменением категориальной неопределенности, которая управлялась через расширение или сужение границ категорий.
Однако исследования функций височной коры позволяют усомниться в непричастности височной доли к процессам категоризации, как утверждают некоторые исследователи (Freedman et al., 2003). Речь идет о веретенообразной извилине (Fusiform G.) – части височной доли в поле Бродмана 37 (BA37), расположенной между окципито-темпоральной и парагиппокампальной извилинами. Основные ее функции – обработка зрительной информации: узнавание лица, тела человека, животных, а также неживых объектов, например, типов автомобилей, а также их зрительного отображения в форме рисунков, картин или скульптур. Веретенообразная извилина принимает участие и в узнавании зрительно предъявляемых слов и чисел (Tan et al., 2011). Опознание лица связывают с работой той части Fusiform G., которая получила название Fusiform face area (FFA). Благодаря этому механизму информация о лице человека, о его состоянии, отношении к другому извлекается в течение нескольких секунд, что очень важно для социального общения. Нарушение FFA ведет к слепоте в отношении лиц (prosopagnosia). У детей с аутизмом активность в этой зоне мозга снижена. Многие полагают, что опознание различных объектов тесно связано с их категориальной идентификацией, которая является одной из важных функций Fusiform G. (McCarthy et al., 1997). Считается, что левая и правая Fusiform G. выполняют разные функции. Левая узнает в объектах черты, сходные с лицом человека, которые могут и не быть реальным лицом. Правая определяет, является ли объект, сходный с лицом, действительно реальным лицом. Таким образом, в выявлении различений в сходных объектах, принадлежащих общей ментальной категории, принимает участие группа нейронов правой, а в отнесении объектов к общей категории – нейроны левой Fusiform G. Из этого следует, что выявление сходства и различия – это разные механизмы.
Хотя взгляды исследователей расходятся относительно зон коры, включенных в процессы категоризации, в том числе и относительно функций височной коры, все они говорят о существовании нейронных сетей, обеспечивающих связь корковых зон со стриопаллидарной системой, в которой роль связующего звена выполняет таламус.
Так, авторы (Grinband et al., 2006), исследовавшие механизмы формирования категориальной неопределенности, не только выявили причастность фронтальной коры к процессам категоризации, но и показали существование сложной фронто-стриатной-таламической сети, включающей медиальную фронтальную извилину, переднюю инсулу, вентральный стриатум и дорзо-медиальный таламус, которая реагировала активностью на категориальную неопределенность.
Связь височных отделов коры высокого уровня со стриатумом выявлена при исследовании семантической категоризации. Методом функциональной магнитно-резонансной томографии (фМРТ) были исследованы мозговые механизмы усвоения второго, не родного – английского – языка у детей в возрасте около 10 лет, родной язык которых китайский, и которые начали изучать иностранный язык в возрасте 6 лет (Tan et al., 2011). Авторы выявили факт взаимодействия активности в Fusiform G. и n. caudate (хвостатом ядре) при решении задачи, требующей отличать английские слова от искусственных слов, в обеих структурах возникала циркуляция мозговой активности. Положительная корреляция была выявлена между уровнями активности в левом хвостатом ядре (ХЯ) и в левой части Fusiform G., а также между их активностью и приобретенным навыком к чтению на английском языке. Активность в двух вышеуказанных структурах мозга авторы рассматривают в качестве нейробиологических маркеров успешности в овладении новым языком. Хотя активность в этих структурах была такой же высокой и при выполнении аналогичной задачи на родном языке, корреляция их активности и ее связь с успешностью чтения на родном языке не обнаружена. Полученные результаты согласуются с более ранними работами, показавшими, что левое ХЯ вовлечено в процессы обработки второго языка – французского (Klein et al., 2006) и немецкого (Rüschemeyer et al., 2005). Показано, что ХЯ играет критическую роль в мониторинге и контроле речевого общения у людей, владеющих двумя языками (Crinion et al., 2006). Так, у пациента с нарушениями в ХЯ, владеющего тремя языками, обнаружено непроизвольное переключение с одного языка на другой (Abutalebi et al., 2000). С другой стороны, Fusiform G., являясь посредником в усвоении второго языка, находится спереди от областей V4 и ТЕО, которые известны как эффективные фильтры для нерелевантной информации. Экстраокципитальная кора включена в процессы управления зрительным вниманием, она выбирает и устраняет среди множества конкурирующих объектов ненужные. Это показано в экспериментах с регистрацией активности нейронов и с применением методов визуализации, отображающей активность мозга (Desimone, Duncan, 1995; Reynolds, Chelazzi, Desimone, 1999; Kastner, 1998; Beck, Kastner, 2005). Такой механизм нужен при использовании иностранного языка. Он должен тормозить лексические процессы, которые связаны с родным языком и мешают использованию иностранного языка. Однако проверка гипотезы, что экстраокципитальная кора также вовлечена при обучении иностранному языку, не подтвердилась. Только циркуляция активности между Fusiform G. и ХЯ является показателем обучения, и их активность в этих структурах предсказывает успешность усвоения второго – иностранного – языка.
Исследование зрительной кортико-стриатной и моторно-стриатной петель показало, что из-за особенностей анатомических связей между корой и стриатумом последний не способен формировать категории (Seger, 2008). Категории создаются на корковом уровне и используются стриатумом и базальными ганглиями для отбора сенсорных категорий и соединения их с категориями моторных реакций (Morris et al., 2003; Seger, Cincotta, 2005).
Другой аспект проблемы категоризации связан с изучением того, как мозг выбирает действие или поведение в целом, которое привязывается к объекту, и какие механизмы используются для обобщения, категоризации моторных реакций. Литературные данные связывают эти функции с базальными ганглиями, тесно взаимодействующими с моторными зонами коры (Frank, 2005; 2006).
Таким образом, многие экспериментальные данные дают основание связывать процесс категоризации с взаимодействием неокортекса со многими подкорковыми структурами, в том числе со стриопаллидарной системой, ее базальными ганглиями. Для проведения исследований такого рода обычно требуется регистрация нейронной активности мозга у животных или использование томографических методов для отображения активности в структурах мозга человека (фМРТ, ПЭТ).
Однако традиционные методы анализа ЭЭГ человека по существу не позволяют разделить активность подкорковых и корковых структур мозга. Эта проблема не решается и с помощью факторного анализа: впервые она разрешается в настоящем исследовании с помощью авторского метода «микроструктурный анализ осцилляторной активности мозга» (МАО), базирующегося на пейсмекерной гипотезе ритмогенеза (Данилова, Дмитриева, 2002; Данилова и др., 2002; Данилова, Быкова, 2003; Данилова, 2009; Данилова, Страбыкина, 2011).
Метод опирается на многочисленные экспериментальные данные, свидетельствующие о связи осцилляций биологических систем с процессами обработки информации. Существуют две альтернативные гипотезы относительно механизмов генерации осцилляторной активности мозга: ансамблевая и пейсмекерная. Согласно первой, осцилляторная активность возникает в ансамбле нейронов за счет обратных и реципрокных связей между его нейронами. Согласно пейсмекерной концепции, существуют нейроны-пейсмекеры, работающие как внутриклеточные генераторы, эндогенная активность которых и определяет появление волновой активности в мозге (Греченко, Соколов, 1979; Соколов, 2003; Греченко, 2008).
Современные исследования нейронной активности свидетельствуют в пользу пейсмекерной концепции ритмогенеза. Экспериментальные данные позволяют утверждать, что биологические системы разного уровня говорят на двух языках: на языке волновой активности и на языке потенциалов действия (ПД). При этом ПД – это в большей степени исполнительная часть диалога.
Сегодня интенсивно исследуются осцилляции на клеточном, молекулярном уровне. Введены понятия генетических осцилляторов, определяющие последовательность и скорость созревания органов у эмбриона, кальциевых осцилляторов, регулирующих концентрацию кальция в отдельном нейроне – показатель его подпороговой активности, предшествующей появлению потенциалов действия нейрона. Сигнал, приходящий к пейсмекерному нейрону, первично вызывает его осцилляции, которые являются подпороговыми для ПД. При увеличении амплитуды волновой активности на определенной ее фазе уже вторично возникает ПД (Pedroarena, Llinas, 1997; Llinas, 2009). Именно осцилляции создают связи между нейронами, используя клеточные механизмы управления частотой и фазой.
Осцилляторная активность выявлена в самых различных биологических системах: у цианобактерий, одноклеточных парамеций, у полностью изолированных нейронов моллюска, у нейронов животных и человека в самых различных структурах мозга (Singer and Gray, 1995; Buzsaki, 2006; Kazantsev et al., 2004; Греченко, 2011; Греченко, 2012).
Широко используется понятие «частотного спаривания осцилляций» (coupling), когда в одной структуре низкочастотные осцилляции через фазу управляют периодами активности высокочастотных осцилляций (Kamarajan et al., 2004; Miskovic et al., 2010).
«Структурное спаривание» означает, что связи осцилляций разной частоты возникают между разными структурами мозга, а не в одном локусе.
Сегодня многие исследователи связывают мозговые механизмы когнитивной деятельности с функциями волновой, осцилляторной активности мозга, рассматривая осцилляции в качестве базового механизма, управляющего когнитивными процессами. Эта позиция получает убедительное подтверждение при изучении нарушений мозговых функций (болезни Альцгеймера, Паркинсона, шизофрении, алкогольной зависимости и др.). Показано, что у пациента с болезнью Альцгеймера нарушена синхронизация тета-осцилляций со стимулом. Ее восстановление произошло после холинергической фармакотерапии, которая улучшала фазовую привязку осцилляций к стимулу и тем самым восстанавливала когнитивную деятельность пациента (Yener et al., 2007). Современная наука позволяет исследовать отношения между генами, осцилляциями и когнитивной деятельностью человека. Исследована генетическая природа тета-осцилляций во фронтальных отведениях, в составе Р300 во время когнитивной нагрузки. Авторы пришли к выводу, что ген холинергического мускаринового рецептора (CHRM2) является лучшим кандидатом на управление тета-осцилляциями во фронтальной области мозга, у алкоголиков его функция нарушена (Begleiter, Porjesz, 2006).
Авторский «метод микроструктурного анализа осцилляторной активности мозга» (МАО), базирующийся на пейсмекерной гипотезе ритмогенеза, позволяет уйти от традиционно используемых суммарных показателей основных ритмов ЭЭГ и обратить внимание на их отдельные частотные составляющие. Он оперирует понятиями частотно-селективных генераторов и выявляет их локальную активность в корковых и подкорковых структурах мозга. Осцилляции, на которых работают частотно-селективные генераторы, извлекаются из состава событийно связанных потенциалов (ССП), полученных при многоканальной регистрации ЭЭГ. Используется модель эквивалентных подвижных токовых диполей для узкополосных частотных составляющих ССП. Координаты эквивалентных диполей частотно-селективных генераторов пересчитываются в координаты Стереотаксического атласа мозга человека (Talairach, Tournoux, 1988). Метод МАО выявляет активированные частотно-селективные генераторы и локализует их в структурах мозга с высоким пространственным и временным разрешением (Данилова, Быкова, 2003; Данилова, Лукьянчикова, 2008; Данилова, 2009; Данилова, Страбыкина, 2011; 2012). Применение метода подтвердило дискретный характер активности частотно-селективных генераторов во времени и пространстве. Выявлена независимость поведения частотно-селективных генераторов, различающихся частотой осцилляции в 1 Гц (Данилова, 2006).
В настоящей работе исследовались мозговые механизмы процессов семантической категоризации в экспериментах, в которых испытуемые решали задачи на категоризацию зрительно предъявляемых слов, принадлежащих категориям «животные», «предметы» и «действия». Отнесение стимулов к двум первым категориям различалось выполнением моторной реакции. При работе с категорией «действия» моторная реакция не выполнялась. В работе представлены результаты исследования только для двух категорий стимулов: «животные» и «предметы», которых объединяет наличие моторных реакций как поведенческого показателя принятия решения.
Процедура и методы исследования
Проведенная работа по своему характеру является психофизиологической – соответственно, экспериментальная процедура имеет психологическую и физиологическую составляющие. Психологическая часть исследования базировалось на разработках, проведенных ранее в лаборатории Т. Н. Ушаковой в Институте психологии РАН (Ушакова, 2012; Ушакова, Павлова, Зачесова, 1989), направленных на изучение когнитивной структуры процесса категоризации с использованием вербальных сигналов. Подготовленный в этих экспериментах вербальный материал для трех категорий стимулов (животные, предметы, действия) учитывал равенство словесных сигналов как физических раздражителей, длительность используемых слов, количество слогов, положение в слове ударного слога и другие их физические особенности. Он и был использован в настоящем исследовании.
Исследование выполнено на 10 испытуемых в возрасте 22–25 лет, которым зрительно на экране компьютера в случайном порядке предъявлялись слова, относящиеся к трем категориям: «животные», «предметы» и «действия». В каждую категорию входило по 10 различных слов, т. е. всего было 30 слов. Каждое слово повторялось восемь раз, т. е. в эксперименте испытуемому предъявлялось 240 слов, принадлежащих трем категориям. Испытуемый получал задание как можно быстрее относить каждое слово к одной из трех категорий, реагируя на слова, принадлежащие домену «животные», одной моторной реакцией (нажатием на клавишу), а на слова, относящиеся к категории «предметы» – двумя последовательными нажимами. Слова, обозначающие «действие», не требовали моторной реакции. Время экспозиции каждого слова – 1 с, интервал между предъявлением слов – 2 с. Программа проведения эксперимента была компьютеризирована на кафедре психофизиологии факультета психологии МГУ имени М. В. Ломоносова, включая сценарий предъявления стимулов с различными метками для каждой категории, которые были синхронизированы с записью ЭЭГ. В специальном протоколе измерялось время каждой моторной реакции. Во время эксперимента производилась запись ЭЭГ с помощью компьютерной системы «Brainsys» фирмы НМФ «Статокин» (Россия). Использовалась 15-канальная регистрация ЭЭГ по международной системе 10–20 % с отведениями в О2, О1, Р4, РЗ, С4, СЗ, CZ, Т6, Т5, Т4, ТЗ, F4, F3, F8, F7. Референтом служил объединенный ушной электрод. Частота оцифровки ЭЭГ 400 Гц. Полоса пропускания 0,3–80 Гц. Применялся режекторный фильтр на частоте 50 Гц. Так как слова каждой категории предъявлялись 80 раз, это позволяло путем усреднения получать событийно связанные потенциалы (ССП). Для каждого испытуемого было получено по три ССП, отдельно для каждой категории слов. Далее полученные ССП обрабатывались программами авторского «метода микроструктурного анализа осцилляторной активности мозга» (Данилова, 2005; 2009; Danilova, Strabykina, 2010; Данилова, Страбыкина, 2011). В программе Brainloc (модель одного подвижного диполя) для отфильтрованных из ССП осцилляций рассчитывались координаты эквивалентных диполей. Поиски дипольных источников производились каждые 2,5 мс. Координаты эквивалентных диполей, рассчитанные в программе BL6 (Ю. М. Коптелов), пересчитывались в координаты Стереотаксического атласа мозга человека (Talairach, Tournoux, 1988).
В работе использован новый электроэнцефалографический показатель активности локальных нейронных сетей – активированные частотно-селективные генераторы, отображающие активность пейсмекерных нейронов (Данилова, Быкова, 2003). Для измерения активности частотно-селективных генераторов подсчитывалась сумма их эквивалентных диполей (Данилова, 2009). При изучении мозговых механизмов процессов семантической категоризации основное внимание было направлено на активность и локализацию частотноселективных тета-генераторов, тесно связанных с процессами памяти и обучения. Для статистики выявления ведущих локусов активности в структурах мозга по показателям активности частотно-селективных генераторов использован метод дисперсионного анализа с повторениями (ДА), примененный к координатам их дипольных источников.
Результаты исследования и их обсуждение
Для группы испытуемых в 10 человек показано статистически достоверное различие в латентных периодах моторных реакций при отнесении слов к категориям «животные» и «предметы». Категоризация слов – названий животных, происходило быстрее, чем слов, относимых к категории «предметы». Среднее значение латентного периода моторных реакций при отнесении к категории «животные» и «предметы» соответственно составили 872,8 мс и 913,6 мс. Достоверность различия латентных периодов моторных реакций подтверждена Т-критерием Вилкоксона (Тэмп = 4,5, Ткрит = 5 при р < 0,01). Полученные результаты согласуются с литературными данными о различии категоризации названий одушевленных и неодушевленных объектов (Ушакова и др., 1989; Kiefer, 2005; Kalenine, Bonthoux, 2007; Марченко, 2010). Однако по числу правильных ответов, уровень которых был высок в нашем исследовании, при отнесении слов к одной из двух категорий группового различия не выявлено (расчет по G-критерию знаков показал недостоверность различия).
Сам факт более быстрой моторной реакции на слова, относящиеся к домену «животные», в сравнении с моторными реакциями на слова, принадлежащие домену «предметы», предполагает, что процесс категоризации предметов является более сложной задачей, требующей больших усилий и энергетических затрат. Для проверки этой гипотезы были измерены уровни активности частотно-селективных генераторов, работающих в разных частотных диапазонах. Уровень их активности измерялся суммой дипольных источников активированных частотно-селективных генераторов, локализованных в различных структурах мозга, т. е. в объеме целого мозга отдельно для каждого частотного диапазона: дельта (1–4 Гц), тета (4–8 Гц), альфа (8–14 Гц), бета (14–30 Гц), гамма1 (30–45 Гц) и гамма2 (55–80 Гц). Активированные частотно-селективные генераторы извлекались из состава индивидуальных ССП, полученных для усредненных реакций ЭЭГ раздельно для категорий «животные» и «предметы». В составе ССП анализировалось два временных кванта: до предъявления слова на интервале в 1 с и после его предъявления на интервале той же длительности.
Было исследовано влияние решения задач на категоризацию на уровень активности мозга в составе ССП до и после предъявления слов. Сравнение активности частотноселективных генераторов, измеренной по сумме их дипольных источников после предъявления слов, относящихся к категории «животные» и «предметы», не выявило их различия ни в одном из частотных диапазонов. Различия в уровне мозговой активности были статистически недостоверны. Однако в составе ССП на интервале в 1 с перед предъявлением слов выявлена зависимость уровня мозговой активности от типа определяемой категории. Это различие в активности было обнаружено только для двух частотных диапазонов: для тета- и альфа-частотно-селективных генераторов.
Активность тета-генераторов оказалась более высокой перед предъявлением слов, относящихся к категории «предметы», чем перед словами, принадлежащими категории «животные». Различие статистически достоверно по Т-критерию Вилкоксона (Тэмп = 9, Ткрит = 10 при р < 0,05 и n = 10). Усиление активности тета-генераторов происходило параллельно с подавлением активности альфа-генераторов, т. е. также на интервале в 1 с перед словами, принадлежащими категории «предметы». Усиление активности тета-генераторов – показатель вовлечения процессов памяти: запоминания, сохранения и извлечения информации. Подавление же активности частотно-селективных альфа-генераторов является другим показателем мозговой активности – эквивалентом депрессии альфа-ритма, свидетельствующим об усилении внимания. В настоящей работе он также измерялся суммой дипольных источников. Различие в уровне активности по альфа-генераторам для двух категорий также статистически достоверно по Т-критерию Вилкоксона (Тэмп= 4, Ткрит = 5, при р < 0,01). Полученные результаты о более высокой активности мозга перед предъявлением слов, относящихся к категории «предметы», в сравнении с категоризацией слов, принадлежащих домену «животные», отображены на рис. 1. Показана зависимость уровня активности тета- и альфа-генераторов от категории на интервалах в 1 с перед предъявлением слов. Только перед стимулами-словами, относящимися к категории «предметы», наблюдался рост мозговой активности, который представлен уменьшением числа дипольных источников альфа-генераторов, что соответствует хорошо известному явлению депрессии альфа-ритма. Подавление альфа-активности развивается параллельно с ростом суммы дипольных источников тета-генераторов, отражающей активацию, связанную с процессами памяти.
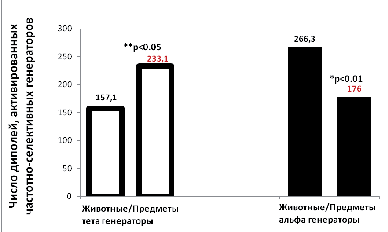
Рис. 1. Различие активации частотно-селективных тета- и альфа-генераторов перед предъявлением слов, относящихся к двум различным категориям: «животные» и «предметы». Перед предъявлением слов, принадлежащих домену «предметы», активность тета-генераторов достоверно выше, чем перед словами, относящимися к категории «животные» (p < 0,05). При этом активность альфа-генераторов перед словами домена «предметы» подавлена в сравнении со словами категории «животные» (p < 0,01)
Таким образом, различие процессов категоризации при работе со словами, принадлежащими разным доменам, получило подтверждение по латентному периоду моторных реакций и уровню мозговой активности, измеренной числом эквивалентных диполей частотно-селективных тета- и альфа-генераторов. Более длинный латентный период моторной реакции, который является поведенческим показателем более трудно решаемой задачи, сочетался с ЭЭГ-показателями активности мозга в виде увеличения суммы дипольных источников для частотно-селективных тета-генераторов и уменьшения числа диполей для альфа-генераторов. Такое изменение активности мозга можно рассматривать как адекватный ответ на требование более сложной задачи, к которой относится выполнение категоризации названий предметов. Однако увеличение активности мозга возникало не после предъявления слов, когда запускался сам процесс категоризации, а перед ним. Этот факт можно объяснить с позиции антиципации, которая мобилизует внимание и формирует готовность отвечать в соответствии с ожидаемой категорией стимулов. Впервые эффект антиципации нами был исследован в опытах с повторением одного и того же звукового стимула, следующего с фиксированным межстимульным интервалом, при инструкции реагировать моторной реакцией на звуковой стимул как можно быстрее (Данилова, Быкова, 2003). Перед предъявлением стимулов возникала вспышка активности частотно-селективных гамма-генераторов, которая усиливала реакцию на звуковой стимул. Локализация в структурах мозга дипольных источников частотно-селективных гамма-генераторов подтвердила это заключение. При выполнении моторной реакции на повторяющийся звук, требовавшем произвольного внимания, активность гамма-генераторов появлялась на интервале
100 мс до стимула: сначала во фронтальной коре, потом смещалась в слуховую кору. Эта же временная и пространственная динамика диполей воспроизводилась и на интервале 100 мс после стимула. При этом по числу диполей после стимула она была более сильной. В индифферентной серии при пассивном прослушивании звуков, не требовавшем моторной реакции, реакция антиципации также присутствовала, но была слабее и локализовалась только в слуховой коре, также как и сенсорный компонент реакции на интервале 100 мс после стимула (Данилова, Быкова, 2003). Эффект прогнозирования событий описывают многие исследователи (Безденежных, 2004; Александров, 2006). Однако, как правило, об антиципации судят по результатам выполняемого ответа после стимула – по укорочению латентного периода моторной реакции, изменению структуры ССП, снижению порога, т. е. по косвенным показателям. В нашем исследовании антиципация возникала в составе ССП как активация частотно-селективных генераторов, появляющаяся до стимула и предсказывающая локализацию активности в тех же структурах мозга после стимула.
В другом исследовании (Данилова, Страбыкина, Плигина, 2012) с запоминанием чисел и опознанием среди них целевых показано, что активность бетаи гамма-генераторов перед нецелевыми (дифференцировочными) стимулами достоверно выше, чем перед целевыми стимулами. Эти результаты также подтверждают факт антиципации, которая позволяет предвидеть появление стимулов определенной категории, а также и то, что в случае ожидания нецелевых стимулов мозг реагирует большей активностью. На этом основании был сделан вывод, что опознание нецелевых стимулов является более сложной задачей, чем опознание целевых.
В настоящем исследовании предъявлялось не две, а три категории стимулов, имеющих равную вероятность. Это создавало большие трудности для формирования антиципации, чем в исследованиях, о которых было сказано выше. Тем не менее, эффект предвидения проявился и в настоящем исследовании. Судя по более длительному латентному периоду моторной реакции, выполнение задачи на категоризацию предметов было более трудным. Испытуемые, ожидая появление слов, принадлежащих категории «предметы», и предвидя большую трудность решения задачи на категоризацию, реагировали большим усилением активности частотно-селективных тета-генераторов на интервале ССП в 1 с до стимула. Последняя измерялась как сумма дипольных источников генераторов в объеме целого мозга. Также перед предъявлением слов, относящихся к категории «предметы», было обнаружено подавление активности частотно-селективных альфа-генераторов, которое можно рассматривать как эквивалент депрессии альфа-ритма. Таким образом, перед предъявлением слов, принадлежащих категории «предметы», выявлено активное использование частотно-селективных тета-генераторов, отображающих процессы памяти, и подавление активности частотно-селективных альфа-генераторов, указывающее на усиление внимания. Эти результаты подтверждают факт формирования антиципации, которая позволяет предвидеть предъявление слов, относящихся к домену «предметы», категоризация которых является более трудной задачей в сравнении с категоризацией слов, принадлежащих домену «животные». В результате перед предъявлением слов, относимых к категории «предметы», усиливаются процессы внимания и памяти.
Мы не получили подтверждения о различии процессов категоризации для этих двух доменов после предъявления стимула на интервале ССП в 1 с. По уровню активности частотно-селективных генераторов ни для одного из шести частотных диапазонов не было выявлено различия. Поэтому мы сделали следующий шаг, применив метод «микроструктурный анализ осцилляторной активности мозга» (Данилова, 2006; 2009), который позволял определять мозговые структуры, вовлеченные в процесс категоризации, и исследовать их зависимость от типа категорий. Метод по данным локализации активированных частотно-селективных генераторов выявляет локальные зоны активности в коре и подкорковых структурах мозга с высоким пространственным и временным разрешением и производит расчет их координат в соответствии со Стереотаксическим атласом мозга человека (Talairach, Tournoux, 1988).
Локализация определялась для тета-генераторов как причастных к процессам памяти и обучению и, следовательно, к формированию в памяти категорий. Исследовалась локализация дипольных источников частотно-селективных тета-генераторов в структурах мозга во время решения задач на категоризацию, т. е. после предъявления слов. Сравнивались локализации частотно-селективных тета-генераторов в составе ССП, полученные для двух категорий: «животные» и «предметы» в интервале в 1 с после стимула. Ранее отмечалось, что в этом интервале ССП мы не обнаружили различий по уровню суммарной активности частотно-селективных генераторов ни для одного частотного диапазона, включая тета.
Для двух задач на категоризацию различие активности было выявлено в Fusiform G., т. е. в той структуре мозга, которую вместе с хвостатым ядром (ХЯ) связывают с формированием категорий. Активность тета-генераторов в Fusiform G. была выше при отнесении слов к категории «животные». Различие подтверждено двухфакторным дисперсионным анализом (ДА) с повторениями (р = 0,048, n = 10, Fкр = 4,113). Хотя статистического различия активности по полушариям не было получено, однако общая тенденция говорит о преобладании активности тета-генераторов в левой Fusiform G. На слова, обозначающие животных, активность в Fusiform G. височной доли возникала только в левой части структуры, а на слова, относящиеся к категории «предметы», активность тета-генераторов возникала в обоих полушариях, но в левом полушарии она преобладала. Полученные результаты хорошо согласуются с литературными данными, полученными другими методами анализа, о вовлечении левой части Fusiform G. в процессы категоризации (McCarthy et al., 1997; Li et al., 2011). В нашем исследовании при отнесении слов к категории «животные» выявлена и вторая зона активности тетагенераторов. Она возникала в экстраокципитальной коре, в языковой извилине (Lingual G.), в поле Бродмана 19 (ВА19). Активность в Lingual G. также была статистически достоверно выше при отнесении слов к категории «животные» (р = 0,048, n = 10, Fкр = 4,113). Поля 17, 18 и 19 – части зрительной коры, организованные по иерархическому принципу, в которой поле 19 реагирует не на отдельные элементы зрительных объектов, а на их целостность (гештальт). Активность в поле Бродмана 19, возникающая на названия животных, говорит об актуализации в памяти их зрительных образов, что менее выражено при работе со словами, относящимися к домену «предметы». Экстраокципитальная кора включена в процессы управления зрительным вниманием, в частности, она выбирает и устраняет среди множества конкурирующих объектов ненужные (Desimone, Duncan, 1995; Reynolds, Chelazzi, Desimone, 1999; Kastner, 1998; Beck, Kastner, 2005). Тот факт, что активность в Lingual G. (ВА19) представлена частотно-селективными тета-генераторами, подтверждает такую интерпретацию. Тета-ритм является ЭЭГ-показателем обращения к процессам памяти, выполняющим функцию как запоминания, так и извлечения информации из памяти.
Третья зона активности тета-генераторов была выявлена в переднем ядре (AN) таламуса, при этом она также была выше при категоризации слов, относящихся к категории «животные». Достоверность различия активности в этой структуре в зависимости от типа выполняемой категоризации подтверждена ДА (р = 0,035, n = 10, Fкр = 5,188). Согласно литературным данным таламус является промежуточным звеном в обмене активностью между зонами коры и стриапаллидарной системой, в том числе и с ХЯ, что и обеспечивает как сам процесс консолидации, так и ее использование при опознании стимулов, а также при решении задач, требующих отнесения стимулов к категориям. Параллельное появление активности тета-генераторов в Fusiform G., в экстраокципитальной коре (ВА19) и переднем ядре таламуса (AN), которая для домена «животных» в этих структурах была статистически выше, чем для домена «предметы», указывает на сложное взаимодействие коры и подкорковых структур мозга, возникающее при категоризации, которое более выражено при опознании категории «животные» (Данилова, Ушакова, 2013). Полученные результаты подтверждают существование сложной фронто-стриатной-таламической сети, вовлеченной в процесс категоризации (Grinband et al., 2006; Tan et al., 2011).
Таким образом, по нашим данным, механизм категоризации более выражен при работе с доменом «животные». Хотя при решении задач на категоризацию слов, принадлежащих домену «предметы», в тех же структурах мозга активность тета-генераторов также присутствует, но она значительно ниже.
Единственной структурой мозга, которая показала большую активность тета-генераторов при работе со словами, принадлежащими домену «предметы», был мозжечок. В составе ССП, полученном для слов домена «предметы», в интервале в 1 с после их предъявления активность выявлена в задней доле мозжечка, на медиальной стороне, в Posterior Cerebellar Tonsil (миндалевидном образовании). Мозжечок играет важную роль в моторном контроле. Он получает входы от сенсорных систем и вовлечен в некоторые когнитивные функции, такие как эмоциональные реакции. Реакция на слова, принадлежащие домену «предметы», в этой структуре оказалась выше, чем на слова, принадлежащие домену «животные». Различие активности статистически достоверно и подтверждено однофакторным ДА (р = 0,046, n = 10, Fкр = 4,413). При этом уровень активности правого полушария был выше левого. Различие статистически достоверно по данным двухфакторного ДА (р = 0,024, n = 10, Fкр = 4,113). Возможно, что более высокая активность мозжечка при отнесении слов к категории «предметы» объясняется двойной моторной реакцией, которую выполняли на стимул этой категории. Доминирование правого полушария позволяет предположить автоматизацию выполнения моторной реакции.
Таким образом, выявлено различие реакций мозга при категоризации слов, принадлежащих двум доменам: «животные» и «предметы». Это различие обнаружено как перед предъявлением слов, так и после их предъявления. Различие реакций выявилось как по уровню активности частотно-селективных генераторов, так и по локализации их дипольных источников, полученных методом «микроструктурный анализ осцилляторной активности мозга». Данные мозговой активности согласуются с поведенческими показателями – латентным периодом моторных реакций, который оказался меньше при категоризации слов, относящихся к домену «животные».
Полученные результаты хорошо объясняются концепцией о существовании сложной фронто-стриатно-таламической сети, выполняющей функции формирования и актуализации категорий (Grinband et al., 2006; Tan et al., 2011). Результаты локализации активности частотно-селективных тета-генераторов в структурах мозга после предъявления слов для решения задач на категоризацию свидетельствуют, что домен «животные» имеет более тесные связи с фронто-стриатно-таламической системой, участвующей в процессах формирования и актуализации категорий. Одно из объяснений более быстрого решения задач на категоризацию для слов – названий животных – их схожесть и меньшее число объектов в составе самого домена «животные» (Марченко, 2010). Однако в нашем исследовании при категоризации одушевленных объектов (животные) выявлен новый фактор, влияющий на принятие решения. Это более высокий уровень активности в структуре Fusiform G. височной доли и в экстраокципитальной коре, в поле 19 Бродмана, в которых хранятся следы зрительной памяти об объектах. Активация представлена частотно-селективными тетагенераторами и указывает на работу со следами памяти, на актуализацию и считывание информации из долговременной памяти. Яркий зрительный образ объекта облегчает процесс принятия решения и сокращает время нужной моторной реакции. Этого не происходит при работе со словами – названиями предметов. Их зрительный образ мало используется в силу того, что за каждым названием определенного предмета может быть закреплено несколько различных зрительных образов, например, стол кабинетный и стол обеденный. Это должно увеличивать время принятия решения.
Наконец, можно предположить, что в ходе экспериментов с задачей на семантическую категоризацию идет процесс обучения, в результате чего категории начинают опознаваться по малому числу признаков. Это должно ускорять их опознание за счет формирования обобщенного образа объектов. Возможно, что при меньшем числе объектов, образующих домен «животные», категория формируется быстрее и это также определяет более короткий латентный период моторной реакции. Высказанные выше предположения в будущем могут быть проверены путем исследования временной динамики локусов мозговой активности, создаваемых активированными частотно-селективными генераторами.
Представленные результаты получены путем определения пространственных координат активированных частотно-селективных тета-генераторов в структурах мозга, что свидетельствует о высокой эффективности авторского метода микроструктурного анализа осцилляторной активности мозга.
Выводы
1. По латентному периоду моторной реакции выявлено различие в сложности выполнения задач на категоризацию слов, относимых к двум доменам: «животные» и «предметы». Категоризация слов – названий животных по латентному периоду моторной реакции происходила быстрее, чем слов, относимых к категории «предметы», что позволяет рассматривать работу со словами, относимыми к домену «предметы», более сложной.
2. Различие двух задач на категоризацию, выявленное по времени реакции и указывающее на большую сложность отнесения слов к категории «предметы», подтвердилось и уровнем активности частотно-селективных тета- и альфа-генераторов на этапе ожидания стимулов. В составе ССП в интервале в 1 с перед предъявлением слов, относимых к категории «предметы», суммарная активность частотно-селективных тета-генераторов усилена, а альфа-генераторов подавлена, что отражает активацию процессов памяти и внимания. Категоризация слов домена «животные» в составе ССП в том же временном интервале в 1 с перед предъявлением слов выполняется при снижении активности частотно-селективных тета- и увеличении активности альфа-генераторов, что свидетельствует о более низком уровне активности мозга на стадии ожидания слов, относимых к категории «животные».
3. Методом микроструктурного анализа осцилляторной активности мозга, выявляющего локальные зоны активности в структурах мозга, показано принципиальное различие процессов категоризации слов при разделении их на две категории: «предметы» и «животные». По локализации дипольной активности частотно-селективных генераторов в составе ССП после предъявления слов их отнесение к категории «животные» вызывает активность в левой части Fusiform G. височной доли, одной из функций которых является участие в процессах категоризации. Параллельно высокая активность возникает в переднем ядре таламуса как промежуточном звене, связывающем зоны коры со стриапаллидарной системой, а также в экстраокципитальной коре (поле 19 Бродмана). Категоризация слов – названий предметов вызывает высокую активность только в заднем мозжечке (Posterior Cerebellar Tonsil). Можно предположить, что высокая активность в мозжечке при работе со словами этого домена отражает появление дополнительного моторного контроля в ситуации относительной неопределенности, связанной с опознанием и отнесением стимулов к категории «предметы».
4. Процесс категоризации является обобщением, возникающим на основе опознания объекта по ограниченному числу признаков. Эту функцию обобщения связывают с процессами обучения, которые формируют связи между зонами коры, хранящими следы памяти об объектах и их названиях, и стриапаллидарной системой, сохраняющей маркеры категорий, которые поддерживают связь со следами памяти в коре. Полученные результаты о локализации источников активности в структурах мозга после предъявления слов свидетельствуют, что домен «животные» имеет более тесные связи со стриопаллидарной системой, участвующей в процессах категоризации.