Введение
Исследование эволюционных предпосылок самосознания (англ. «self-awareness») является одним из магистральных направлений современной сравнительной психологии и когнитивной этологии, базирующихся сразу на нескольких методических подходах [Gallup, 2020]. В настоящий момент развитие получает точка зрения, согласно которой самосознание является модульным феноменом и каждый из модулей получает свое независимое развитие в филогенезе. Франс Де Вааль высказывает предположение о том, что самосознание, как в фило-, так и в онтогенезе формируется постепенно — слой за слоем [De Waal, 2019].
Существуют три основных подхода к поиску признаков самосознания у животных: 1) изучение способности к самоузнаванию в зеркале [Gallup, 2020]; 2) способность животных различать собственный запах (англ. «olfactory mirror») [Gatti, 2020; Horowitz, 2017]; 3) изучение способности воспринимать свое тело в качестве физического препятствия для решения проблемы (англ. «body-awareness» ) [Dale, 2017; Lenkei, 2020; Lenkei, 2021].
О наличии у субъекта представления о физических свойствах своего тела может свидетельствовать способность спонтанно (без дополнительного обучения) решать задачи, для которых требуется учитывать эти свойства. В настоящее время используют два варианта таких задач. При помощи одного оценивали способность детей, слонов и собак оперировать представлением о том, что их тело имеет вес и понимать, что вес тела может быть препятствием для выполнения экспериментальной задачи [Brownell, 2007; Dale, 2017; Lenkei, 2021]. Второй позволяет оценить способность оперировать представлением о размере своего тела — соотносить его с размером и формой отверстия, через которое субъекту необходимо пройти [Brownell, 2007; Lenkei, 2020].
Достоинством этого подхода является то, что теоретически он применим к широкому кругу видов. Однако при его применении возникает та же проблема, что встает при использовании других методов, направленных на изучение мышления животных — как отличить оперирование представлениями от быстрого обучения решению экспериментальной задачи. Ниже мы рассмотрим имеющиеся к настоящему моменту исследования, посвященные изучению способности животных и человека учитывать собственный вес (англ. «body weight awareness») [Brownell, 2007; Dale, 2017; Lenkei, 2021].
В работе с детьми [Brownell, 2007] в первом тесте стоящего на коврике ребенка просили подкатить к экспериментатору привязанную к этому коврику тележку. Для того чтобы выполнить задачу, ребенку нужно было сойти с коврика (попытку толкать тележку, не сойдя с коврика расценивали как ошибочное действие). Во втором тесте ребенок сидел на коврике и слушал короткую сказку. Когда сказка заканчивалась, экспериментатор просил ребенка подать ему коврик. Дети в возрасте 18 месяцев справлялись с обоими вариантами этой задачи только после одного, а чаще нескольких ошибочных действий. В возрасте 22—26 месяцев число ошибочных действий достоверно снижалось, а некоторые дети решали эти задачи с первой пробы. Эти результаты могут свидетельствовать о том, что представление о свойствах своего тела (а именно о том, что оно имеет вес) на втором году жизни только начинает формироваться.
В работе со слонами [Dale, 2017] животных предварительно обучили по команде подавать экспериментатору палку. Далее слонам предъявили 48 тестовых проб и два типа контрольных проб (тоже по 48 каждого). В начале каждой пробы слона заводили на ковер. В тестовых пробах палка была привязана к ковру. Экспериментатор стоял на таком расстоянии от ковра, что передать ему палку можно было только сойдя с ковра. Контрольные пробы, в которых палка не была привязана к ковру, позволяли выяснить, сходят ли слоны с ковра только в той ситуации, когда это действительно необходимо для решения задачи. Два типа контрольных проб различались только тем, что в одном из них экспериментатор тянул за привязанную к ковру веревку, создавая натяжение ткани под ногами слона. Одной группе животных сначала предъявили все 48 тестовых проб, а затем контрольные. С двумя другими группами эксперимент начинали с тех или иных контрольных проб. Сравнение результатов, как всех 48 проб каждого типа, так и первых 12, показало, что слоны достоверно чаще сходили с ковра в тестовых пробах, в которых это было действительно необходимо. Четыре слона из разных групп не совершили ни одного ошибочного действия в первых 12 пробах. Четыре других допустили только одну ошибку в первых 12 пробах. Двое животных из той группы, у которой первыми были тестовые пробы, в последующих контрольных пробах ни разу не сходили с ковра. Эти результаты говорят о том, что правильное действие в тестовых пробах не было сформировано в результате обучения.
Похожую методику использовали для оценки body awareness у 54 собак [Lenkei, 2021]. В этой работе дополнительно использовали третий тип контрольных проб, в котором палка была привязана к крюку, закрепленном в полу рядом с ковром, что, с одной стороны, не позволяло подать ее экспериментатору, а с другой — не создавало натяжения коврика под ногами, при попытке сделать это. Чтобы уменьшить влияние обучения, каждой собаке предъявили 4 пробы каждого типа (4 тестовые и 12 контрольных), чередуя их в квазислучайном порядке. В тестовых пробах на ковре оставалось меньше 15% животных. Собаки достоверно чаще и быстрее сходили с ковра в тестовых пробах, чем во всех трех типах контрольных. Они сходили с ковра достоверно позже в тех пробах, в которых палка была привязана к крюку, причем чаще делали это, отпустив палку, тогда как в тестовых пробах они чаще сходили с ковра не отпуская палку. В тех контрольных пробах, в которых экспериментатор тянул за веревку, привязанную к ковру, собаки оставались на нем достоверно чаще, чем в тестовых пробах, что свидетельствует о том, что ощущения натяжения ткани под ногами недостаточно для того, чтобы собака сошла с ковра. В целом, эти результаты указывают на то, что собаки понимают структуру этой задачи и обладают представлениями о том, что их тело имеет вес.
Целью настоящего исследования является изучение способности серых крыс учитывать вес собственного тела в качестве препятствия для решения экспериментальной задачи. Крысы являются распространенными модельными объектами для изучения когнитивных процессов [Barnes, 1979; Morris, 1984]. Крысы обладают пространственной эксплицитной памятью и могут ориентироваться на местности с помощью внешних знаков [Treisman, 1980]. Крысы способны выучивать правила решения задачи. Так, в исследовании 2008 года была продемонстрирована способность крыс ориентироваться на последовательности визуальных стимулов [Murphy, 2008]. Между тем данные грызуны ни разу не участвовали в экспериментах, направленных на изучение учета веса собственного тела. Ранее нами было проведено исследование, продемонстрировавшее способность крыс учитывать границы собственного тела [Хватов, 2016; Lenkei, 2021]. В силу специфики вида мы не могли организовать экспериментальную задачу аналогично тому, как это было сделано в исследовании на детях, слонах и собаках [Brownell, 2007; Dale, 2017; Lenkei, 2021]. Таким образом, нами была разработана оригинальная методика для изучения способности крыс учитывать вес собственного тела.
Материалы и методы
Испытуемые: 41 самец крысы Rattus norvegicus, наивные особи в возрасте от 2 до 6 мес. В ходе эксперимента животные содержались в индивидуальных клетках.
Экспериментальная установка представляла собой стеклянный ящик без потолка (площадь 1000х950, высота 500 мм). Внутри ящика с его противоположных сторон параллельно плоскости дна на высоте 300 мм располагалось две полки, занимавших всю ширину ящика и имевших длину 320 мм каждая. Полки были соединены тремя мостиками (50 мм шириной каждый), расположенными параллельно друг другу. Два мостика располагались по краям на расстоянии 200 мм от стенки ящика, третий — в центре на расстоянии 200 мм от каждого из боковых мостиков. Мостики крепились на едином металлическом стержне, располагавшемся точно по центру их длины. Благодаря этому мостики могли фиксироваться в разных позициях (рис. 1).
Незакрепленная позиция: мостики, будучи сбалансированными, располагались параллельно дну, соединяя обе полки. При этом легкое нажатие на один из краев мостика заставляло его смещаться по вертикали.
Закрепленная позиция: мостики дополнительно крепились к полкам с помощью щеколд (со стороны дна), благодаря чему нажатие на них не влекло за собой их смещение.
Эксперимент состоял из двух серий: ознакомительной и экспериментальной. В начале каждой пробы крыса помещалась в центр одной из полок внутри экспериментальной установки (далее — полка № 1). В центре противоположной полки (далее — полка № 2) располагалась приманка (сыр).
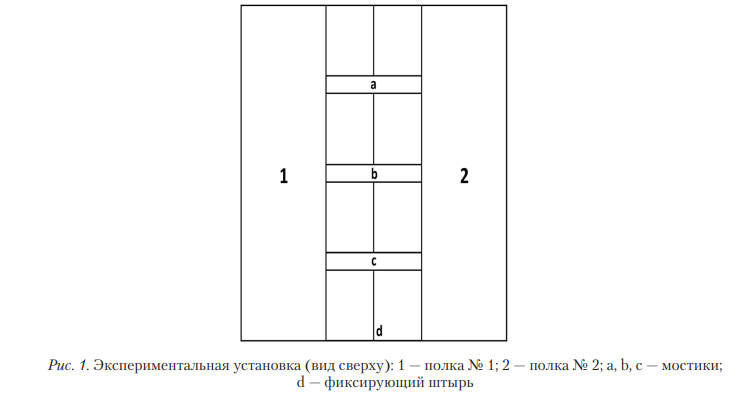
Ознакомительная серия состояла из 27 проб. Все мостики находились в закрепленной позиции. Задачи серии: сформировать у крыс навык достижения приманки, выявить, формируются ли у крыс индивидуальные предпочтения определенного мостика для перехода на полку № 2.
Экспериментальная серия состояла максимум из 36 проб. В каждой пробе только один из трех мостиков находился в закрепленной позиции; расположение этого мостика варьировалось квазислучайно: во-первых, закрепленный мостик 12 раз должен был находиться в каждой позиции (слева, в центре или справа), во-вторых, один и тот же мостик не мог оставаться в закрепленной позиции более двух проб подряд для одной особи. Задача серии: сформировать у крыс навык выбора, подходящего для прохождения мостика. Серия продолжалась либо до достижения крысами критерия обученности — 9 успешных проходов подряд без падений (p=0,001, биномиальный тест), либо максимально продолжалась 36 проб.
В ходе эксперимента фиксировались следующие зависимые переменные.
— Количество успешных проходов по закрепленному мостику с полки № 1 к полке № 2 (в ознакомительной серии).
— Количество попыток прохода по мостику (в экспериментальной серии) — такая попытка могла быть успешной, если крыса проходила по закрепленному мостику, или неуспешной, если крыса падала
— Количество падений с незакрепленных мостиков (в экспериментальной серии).
— Количество «пробующих движений» (в экспериментальной серии). Под пробующими движениями мы понимали такое поведение крысы, при котором она, располагаясь четырьмя лапами на полке № 1, вытягивала морду в направлении мостика. Затем крыса осуществляла нажатие на мостик одной из передних лап. Животное осуществляло несколько таких нажатий в течение примерно 5—10 секунд, после чего действовало в зависимости от результата. В случае если мостик оказывался закреплен, крыса переходила по нему к полке № 2. В случае если мостик не был закреплен, такие пробующие движения крысы приводили к некоторому смещению мостика под давлением лапы крысы: край мостика опускался ниже уровня полки, после чего возвращался в исходную позицию. Описанные пробующие движения обнаруживались в поведении некоторых крыс, начиная со второй пробы экспериментальной серии.
Статистический анализ данных. В ознакомительной серии для выявления предпочтения положения мостика использовался критерий хи-квадрат Пирсона, с помощью которого сопоставлялись эмпирические распределения количества проходов по левому, центральному и правому мостикам, полученные в результате эксперимента, с равномерным распределением (вероятность прохода по каждому мостику 33,3%).
В экспериментальной серии для выявления факторов, влиявших на выбор мостика для осуществления попытки прохода, мы использовали факторный дисперсионный анализ ANOVA. В качестве переменных предикторов использовались: фиксация мостика (закрепленный/незакрепленный), положение мостика (левый/центральный/правый).
Для выявления влияния пробующих движений на количество падений был осуществлен линейный регрессионный анализ, где в качестве переменной-предиктора использовалось количество пробующих движений, в качестве зависимой переменной — количество падений.
Все математические расчеты осуществлялись в Statsoft Statistica (версия 10.0.1011.0).
Результаты
Результаты ознакомительной серии. Каждая из крыс успешно достигала приманку, переходя от полки № 1 к полке № 2, в каждой из проб. У всех крыс было выявлено индивидуальное предпочтение центрального мостика (по сумме за 27 проб) — критерий х2 (df=2; p=0,001). Суммарно крысы прошли по левому мостику 146 раз, по центральному — 780, по правому — 154 (х2= 327,803; df=2; p=0,001).
По результатам экспериментальной серии крысы разделились на 3 группы.
1-я группа крыс — 14 особей, достигнувшие критерия обученности. Крысы допустили от 2 до 5 падений (M=3,07; SD=1,07). У этих крыс обнаруживаются пробующие движения после нескольких падений, начиная со 2-й пробы: перед попыткой, прохода по мостику, крысы данной группы осуществляли пробующие движений и далее действовали в зависимости от результата.
2-я группа крыс — 8 особей, не достигшие критерия обученности за 36 проб. Крысы допустили от 20 до 27 падений (M=23,00; SD=2,58). У этих животных было выявлено 5 пробующих движений.
3-я группа крыс — 19 особей, совершивших от 2 до 5 падений (S=2,89; SD=0,80) и далее не совершавших попыток прохождения по мостику, оставаясь на полке № 1. После совершенных падений в последующих пробах крысы, будучи помещенными на полку № 1, либо вообще не подходили к мостикам, либо подходили к ним, совершали пробующие движения, но далее не шли.
Единственным предиктором, влиявшим на осуществление попытки прохода по мостику у 1-й группы крыс, являлась фиксация мостика: крысы достоверно чаще осуществляли попытки прохода по закрепленному мостику (N = 14; F(1, 78) = 290,278; p = 0,00001). Предиктор положения мостика (левый/центральный/правый) не оказывал влияния, также как и его взаимодействие с предиктором фиксации мостика (табл. 1, рис. 2).
Таблица 1
Результаты 14 крыс 1-й группы: оценка влияния различных предикторов
(фиксации мостика, положения мостика) на количество попыток прохода
по мостикам — факторный ANOVA
|
Предикторы |
SS |
sd |
MS |
F |
Р |
|
Фиксация мостика |
84,0000 |
1 |
84,0000 |
290,278 |
0,00001 |
|
Положение мостика |
0,5000 |
2 |
0,2500 |
0,864 |
0,425498 |
|
Фиксация мостика вместе с положением мостика |
0,9286 |
2 |
0,4643 |
1,604 |
0,207566 |
F(1, 78)=290,28, p=0.0001
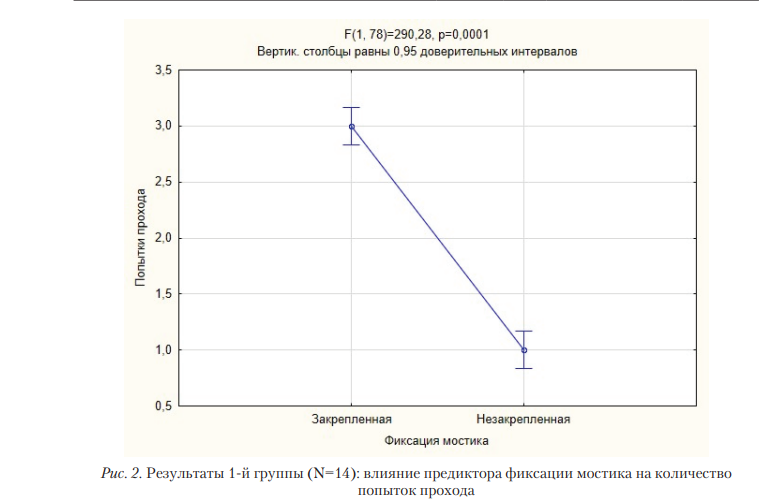
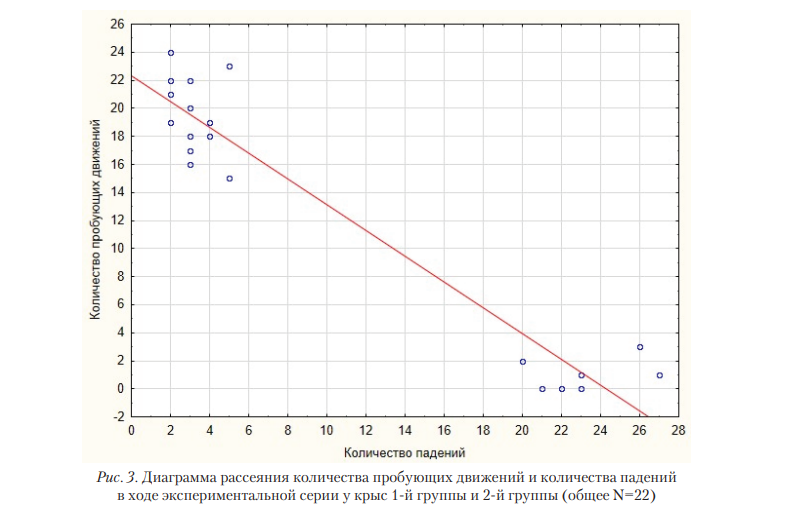
В целом, за все пробы экспериментальной серии крысы из 1-й группы допустили 43 падения, и совершили 275 пробующих движений, крысы из 2-й группы допустили 161 падение и осуществили 5 пробующих движений. Регрессионный анализ показал наличие отрицательной связи между количеством пробующих движений и количеством падений (R=0,963; B=-1,008; p=0,0001) (рис. 3).
Обсуждение результатов
Мы полагаем, что в ознакомительной серии предпочтение всеми крысами центрального мостика для передвижения объясняется тем, что через него пролегает кратчайший путь к приманке. Этот фактор является побочным, однако он исключался квазислучайным положением закрепленного мостика во второй серии. Выделение 3-й группы крыс в экспериментальной серии мы объясняем тем, что для этих животных падения являлись сильным стрессом, в связи с чем далее они не решались совершать попытки продвижения по мостикам.
Вероятно, это обусловлено типом нервной системы крыс. Между тем на основе результатов 14 особей из первой группы мы можем констатировать, что серая крыса способна учитывать вес собственного тела при взаимодействии с объектами окружающей среды. Мы полагаем, что именно в ходе выявленных «пробующих движений» крысы осуществляли сопоставление веса собственного тела с прочностью опоры (закрепленным или незакрепленным мостиком).
При этом, как и в эксперименте со слонами [Dale, 2017], детьми [Brownell, 2007] и собаками [Lenkei, 2021], для крыс их собственное тело сначала являлось препятствием для решения задачи, но затем они стали использовать его в качестве средства для подбора подходящего мостика. Крысы первой группы выучили правило: перед осуществлением прохода по мостику необходимо проверить его на прочность. При этом важно отметить, что данное действие осуществлялось не механически (как процедурный навык — вроде нажимания кнопки или педали) — животное оценивало результат пробующего движения и осуществляло проход только в случае, если после нескольких нажатий мостик не проваливался. Подчеркнем, что при нажатии в ходе поисковых движений на незакрепленный мостик последний не обрушивался, а лишь немного опускался, затем возвращаясь в исходную позиции. Соответственно, крысы делали вывод о его прочности на основании этих признаков. Полученные в ходе настоящего исследования данные согласуются с установленным ранее фактом способности серых крыс принимать в расчет границы собственного тела при проникновении в отверстия различных размеров [Хватов, 2016].
Таким образом, в нашем исследовании была продемонстрирована возможность экспериментального выявления способности к учету веса собственного тела у крыс. К настоящему моменту признаки способности учитывать вес собственного тела обнаружены у детей в возрасте 22—26 месяцев [Brownell, 2007], слонов [Dale, 2017] и собак [Lenkei, 2021]. Использованная нами методика расширяет набор тестов, применимых для исследования многоаспектного феномена восприятия животными собственного тела [De Waal, 2019]. Она может быть применена для широкого круга видов, что позволит проследить развитие этой когнитивной способности в филогенезе.