Введение
Процессы формирования ассоциативных связей занимают центральное место в теории обучения (Резникова, 2005; Reznikova, 2007; Полетаева и др., 2017), и до сих пор в этой области можно ожидать открытия новых форм обучения (Reznikova, 2012). Расширение спектра исследуемых видов позволяет достичь принципиально новых результатов. Так, использование столь оригинальной экспериментальной модели, как избегание опасности жужелицами, позволило описать новую форму обучения — «обучение из каталога» (Reznikova, Dorosheva, 2013). Изучение формирования ассоциативных связей с использованием стимулов различной модальности и при разных режимах и способах обучения является одним из наиболее актуальных вопросов современной сравнительной психологии, затрагивающим особенности научения, мышления и памяти у различных видов животных и человека. Большинство современных исследований в этой области проводится на мышах и крысах разных генетических линий, что позволяет манипулировать точечными мутациями и получать все более детальные представления о генетических и молекулярных механизмах способностей к обучению (Перепелкина и др., 2011; Голибродо и др., 2014; Matzel et al., 2011). По нашему мнению, для достижения баланса между познанием тонких механизмов формирования памяти и пониманием общей картины когнитивных процессов необходимы сравнительные исследования на разных видах. Так, использование процедуры выбора подкрепляемых визуальных стимулов в экспериментах с дикими полевыми мышами Apodemus agrarius выявило необычно высокие способности этого вида к различению количества предметов, сравнимые с высшими приматами (Vorobyeva et al., 2013) и неизвестные для лабораторных линий на основе Mus musculus.
Исследования влияния режимов обучения на успешность формирования ассоциативных связей уходят корнями в ранние этапы становления сравнительной психологии. Впервые вопрос о связи между режимом и успешностью обучения был затронут в классических психологических работах Г. Эббингауза (Ebbinghaus, 1885/1964) и А. Йоста (Jost, 1897), которые пришли к выводу о целесообразности распределения повторений во времени для лучшего запоминания. Этот феномен получил название «the spacing effect for memory» и впоследствии был связан с формированием долговременной (или консолидированной) памяти (Atkinson, Shiffrin, 1968). В эмпирических психологических исследованиях на людях, с предъявлением как вербальных, так и моторных задач, было показано, что при изменении продолжительности сеансов обучения и интервалов между ними оптимальный для запоминания период имеет значительную вариативность при различных задачах (Фресс, Пиаже, 1966; обзор см: Cepeda et al., 2006). В последние десятилетия накопились многочисленные данные, полученные на самых разных объектах, от моллюсков и насекомых до грызунов и приматов, включая людей, свидетельствующие о значительной вариабельности межтестовых интервалов (МТИ) между сеансами обучения, оптимальных для формирования ассоциаций (обзор см: Smolen et al., 2016), а само понятие о долговременной памяти претерпело значительные изменения (обзор см : Григорьян, Маркевич, 2014). Поэтому при работе с различными видами поиск оптимальных МТИ является актуальной задачей для понимания процессов формирования долговременной памяти и управления вниманием.
Отметим, что в русскоязычной литературе употребляются термины «концентрированное» или «интенсивное» обучение (massed training) и «разнесенное» или «расставленное» обучение (spaced training) (Воробьева и др., 2016). Мы в изложении результатов своих экспериментов используем более конкретные термины: «короткие/длительные МТИ».
В современных исследованиях на животных, посвященных различиям в эффективности обучения при использовании разных интервалов между предъявлением стимулов, можно выделить две основные экспериментальные парадигмы. Первую можно назвать «нейрофизиологической». В нейрофизиологических работах в качестве экспериментальной схемы используется, главным образом, формирование ассоциативной связи между условным стимулом (например, звуковым сигналом) и безусловным стимулом (например, электро- кожным раздражением) (обзор см: Воробьева и др., 2016). В этой парадигме исследуется поведение животных в разных режимах и при действии различных внешних и внутренних факторов, при отсутствии для них возможности выбирать стимул. Большинство данных, полученных как на позвоночных, так и на беспозвоночных животных, говорит о том, что для формирования долговременной памяти более благоприятны длительные МТИ, однако стоит заметить, что понятие «длительности» существенно варьирует в разных экспериментах (обзоры см: Philips et al., 2013; Smolen et al., 2016). Вторую экспериментальную парадигму можно назвать «когнитивной»: она связана с формированием ассоциативных связей в ситуациях выбора между подкрепляемым и не подкрепляемым стимулом и предоставляет возможность постановки когнитивных задач (обзоры: Резникова, 2011, 2015). Для сравнения с психической деятельностью человека особенно важно исследование мысленного установления связей между частями задачи, которое может происходить у некоторых видов животных при разных режимах обучения (Зорина и др., 2013). «Когнитивная» парадигма используется в таком актуальном и быстро развивающемся направлении, как оценка изменений внимания и памяти, связанных с возрастом и с различными ментальными нарушениями (Matzel et al., 2011; Barak et al., 2013; Wohr, Scattoni, 2013).
Мы использовали «когнитивную парадигму» для выявления особенностей различения и запоминания визуальных стимулов при разных режимах обучения у джунгарского хомячка дикого типа, ранее не исследованного в этом плане. Полученные данные позволяют судить о некоторых особенностях когнитивной деятельности у этого вида и предложить его в качестве одной из экспериментальных моделей для исследования влияния режимов обучения на успешность формирования ассоциативных связей.
Материалы и методы
В экспериментах участвовали джунгарские хомячки (9 самцов и 1 самка). Варьировали визуальные стимулы и режимы обучения (с короткими и с длинными МТИ). Хомячки были потомками животных, в 1980-е гг. отловленных в Карасукском районе Новосибирской области. Они содержались при естественном освещении и свободном доступе к воде и корму, в индивидуальных клетках с подстилкой из древесной стружки. За сутки до начала тестов у животных убирали корм. За 20—40 минут до начала теста животных в их индивидуальных клетках переносили в звукоизолированную комнату. Тесты проводили с 10:00 до 17:00. Эксперименты проведены с соблюдением правил проведения научных исследований с использованием экспериментальных животных, утвержденных распоряжением Президиума АН СССР от 2 апреля 1980 года 12000 — № 496 и приказом Минвуза СССР от 13 сентября 1984 года № 22.
Экспериментальная установка представляла собой круглую арену диаметром 33 см, на стенках которой закрепляли 2 контейнера (12x12x2 см) со сменными дверцами из плотной белой бумаги. На поверхность дверцы наносились подкрепляемые визуальные стимулы («пятна») произвольной формы (рис. 1). Оценивали, насколько успешно животное научалось связывать подкрепляемый стимул и вознаграждение. Отметим, что в статье термин «обучение» используется для обозначения процесса, а «научение» — для обозначения результата (см также: Резникова, 2005).
В основе наших экспериментов лежит классическая методика дифференцировки стимулов (см.: Зорина и др., 1989; Зорина, Полетаева, 2002). «Экспериментом» (рис. 2 А ) мы называем совокупность тестов, в ходе которых животные в одном режиме решали одну задачу (например, «эксперимент с короткими МТИ, выбор «пятно—нет пятна»). За основу дизайна тестирования мы взяли методику, разработанную Г.А. Мазохиным—Поршняковым (1984) для пчел и позднее адаптированную нами для грызунов (Vorobyeva et al., 2013), поскольку эта схема предполагает высокие требования к критерию научения и полностью исключает влияние безусловного стимула (запаха пищи), что особенно важно для экспериментов с грызунами. Согласно этой методике, во всех экспериментах каждый тест включал три стадии: «ознакомление», «обучение» (три повторности) и «экзамен» (рис. 2 Б). Количество экзаменов равно количеству тестов.
На стадиях «ознакомления» и «обучения» внутрь контейнера с подкрепляемым визуальным стимулом, изображенным на дверце, помещали приманку (кусочек грецкого ореха), второй контейнер оставался пустым. На стадии «ознакомления» животному давали возможность открыть оба контейнера (лапами или носом приподнять бумажную дверцу), найти и съесть приманку. На стадии «обучения», если зверек делал правильный выбор (заходил в контейнер с приманкой), то получал вознаграждение и на время подготовки к проведению следующей стадии теста его помещали в домашнюю клетку; если ошибался, то в качестве наказания его помещали на 5 минут в пустой темный контейнер. Даже если зверек успешно находил приманку с первой попытки, с ним все равно проводили три повторности «обучения». Если животное во время любой из стадий тестирования в течение пятнадцати минут отказывалось заходить в какой-либо контейнер, его удаляли с арены и помещали в домашнюю клетку, и в этот день оно в тестировании более не участвовало.
После каждой стадии теста меняли взаиморасположение контейнеров, чтобы исключить возможность выбора по принципу «справа—слева» («предпочитаемая сторона)», а также привыкание животных к внешним ориентирам. Меняли также и дверцы, чтобы исключить возможность ориентации животного по дополнительным стимулам (пятнам, складкам); дверцы, поврежденные зверьками, в дальнейшем не использовались. Перед каждой стадией теста арену и контейнеры протирали спиртом. После стадий «ознакомления» и «обучения» (в трех повторностях) проводился «экзамен» по той же схеме, но без приманки, что iисключало использование запаха пищи при выборе контейнера. Перерывы между «ознакомлением», повторностями «обучения» и «экзаменом» составляли по 5—7 минут. Фиксация поведения проводилась с помощью видеокамеры (SONY DCR-SX44E), экспериментатор находился вне поля зрения животного. Для статистического анализа использовали только результаты экзаменов.
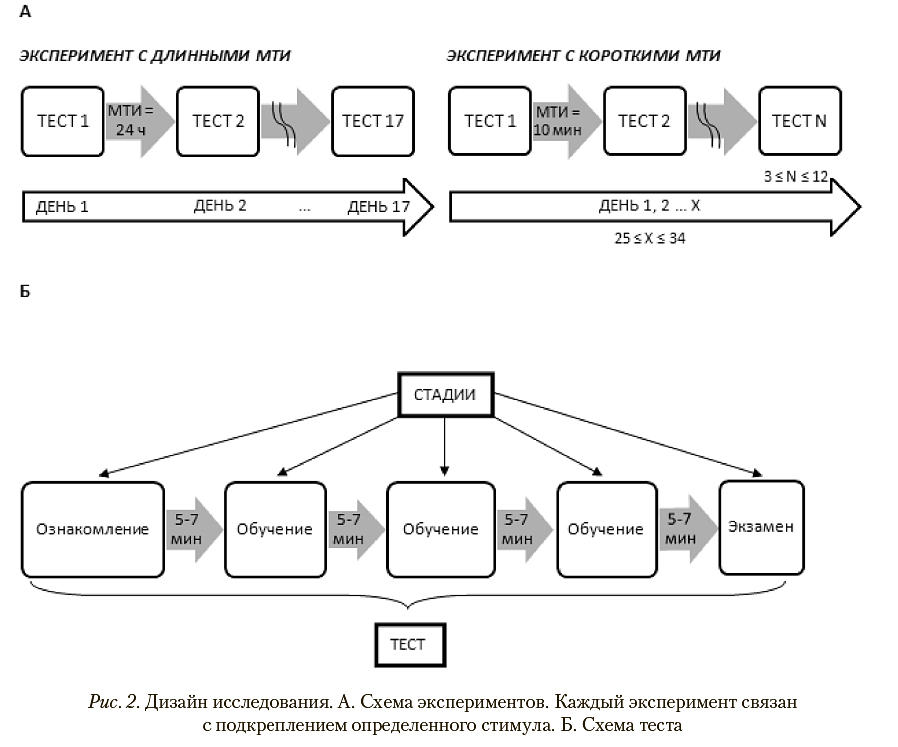
Выбранный нами дизайн тестирования с использованием экзаменов находится в рамках экспериментальной парадигмы вариативного подкрепления, разработанной К. Прайор (Pryor et al., 1964; Pryor, Ramirez, 2014): подкрепляя не каждое проявление нужного поведения, можно не только поддерживать выученное поведение на определенном уровне надежности, но даже и усиливать его. В нашей экспериментальной схеме i |ено.дкретляемые выборы (экзамены) присутствовали во всех трех экспериментах. Исключение всех возможных факторов (ориентация по предпочитаемой стороне, ориентирам вне арены, внешним признакам дверцы, запаховому градиенту в арене) является стандартным приемом избежания превдо- научения (Резникова, 2005). Закономерность предлагалась только одна: за выбор подкрепляемого стимула зверек получал приманку, за выбор второго стимула — наказание. Ранее эта методика применялась нами для оценки индивидуальной вариабельности способностей джунгарских хомячков к различению множеств визуальных стимулов (Роговая и др., 2016).
В первом эксперименте («выбор пятна») подкреплялся выбор контейнера с пятном на крышке (черное пятно 9x5 см на белом фоне). Эксперимент включал две серии тестов: с короткими и с длинными МТИ. В режиме с длинными МТИ 10 животных было протестировано 17 раз, по одному разу в сутки (680 «обучающих сеансов» и «ознакомлений»). Выбор такого количества тестов основан на проведенном ранее исследовании (Роговая и др., 2016) : в нем из числа животных, протестированных в режиме с длинными МТИ 17 раз и не связавших стимул с подкреплением, было случайным образом выбрано 6, и с ними число тестов в том же режиме было доведено до 30. Поскольку ни один зверек при таком количестве тестов показателей обучения не улучшил, мы в данной работе определили количество тестов 17 как достаточное.
В режиме с короткими МТИ было протестировано 9 животных (8 самцов и 1 самка), участвовавших в предыдущей серии тестов (от 900 до 1224 «обучающих сеансов» и «ознакомлений»). Зверьки тестировались ежедневно в течение двух—пяти дней; перерывы между тестами составляли 10 минут. Каждый день тестирование продолжалось до тех пор, пока животное не отказывалось заходить в контейнер в течение 15 минут с момента начала стадии теста; таким образом, зверьки проходили от 3 до 12 тестов в день. Работа с каждым животным продолжалась до достижения научения, т. е. до момента, когда разница между правильными и неправильными выборами в ходе экзаменов начинала значимо отличаться от случайной. Животные, не связавшие стимул с подкреплением за 25 тестов, т. е. выбиравшие контейнер в экзаменах случайным образом, считались не научившимися, но из дальнейших экспериментов не исключались.
Во втором эксперименте («выбор меньшего пятна») пятна были на обеих дверцах. Подкрепляемым стимулом служило черное пятно меньшего размера (5x2 см), а не подкрепляемым — пятно большего размера (9x5 см). 9 животных были протестированы в режиме с короткими МТИ (от 900 до 1008 «обучающих сеансов» и «ознакомлений»).
В контрольном эксперименте выяснялось возможное предпочтение животными правой или левой стороны. Одинаковые пятна помещались на обе крышки, но подкрепление было только в одном из контейнеров. Каждое из 9 животных было протестировано 10 раз в режиме с короткими МТИ.
При статистической обработке данных доли правильных выборов сравнивали со случайным выбором с помощью биномиального критерия. Доли научившихся и не научившихся зверьков сравнивали с помощью точного критерия Фишера с поправкой Бонферрони.
Результаты
В первом эксперименте («есть пятно/нет пятна») в серии тестов с длинными МТИ только один зверек из 10 продемонстрировал научение после 17 тестов (табл. 1).
Таблица 1
Доли выборов подкрепляемого визуального стимула в 17 «экзаменах» у джунгарских хомячков при тестировании с длинными МТИ
|
Номер животного |
Доля правильных выборов |
Значение биномиального критерия, Z |
Уровень значимости |
|
4 |
0,71 |
1,029 |
p>0,05 |
|
8 |
0,63 |
0,532 |
p>0,05 |
|
124* |
0,82 |
1,715 |
p<0,05 |
|
125 |
0,47 |
0 |
p>0,05 |
|
152 |
0,35 |
— 0,686 |
p>0,05 |
|
153 |
0,65 |
0,686 |
p>0,05 |
|
184 |
0,71 |
1,029 |
p>0,05 |
|
211 |
0,47 |
0 |
p>0,05 |
|
255 |
0,63 |
0,532 |
p>0,05 |
|
285 |
0,47 |
0 |
p>0,05 |
Примечание: «*» — особи, научившиеся выбирать подкрепляемый стимул.
В серии тестов с короткими МТИ 7 зверьков из 9 (77,8%) продемонстрировали научение, им потребовалось от 25 до 34 тестов (табл. 2). Доля научившихся зверьков достоверно отличается от доли зверьков, научившихся в серии тестов «есть пятно/нет пятна» с длинными МТИ (точный тест Фишера с поправкой Бонферрони, p=0,05).
Таблица 2
Доля выборов подкрепляемого визуального стимула в 25—34 «экзаменах» у джунгарских хомячков при тестировании с короткими МТИ
|
Номер животного |
Количество тестов |
Доля правильных выборов |
Значение биномиального критерия, Z |
Уровень значимости |
|
4 |
25 |
0,47 |
1,029 |
p>0,05 |
|
8* |
27 |
0,75 |
0,532 |
p>0,05 |
|
124* |
25 |
0,84 |
1,715 |
p<0,05 |
|
125* |
29 |
0,76 |
0 |
p>0,05 |
|
152* |
34 |
0,74 |
— 0,686 |
p>0,05 |
|
153* |
25 |
0,92 |
2,833 |
p<0,05 |
|
184* |
25 |
0,88 |
2,155 |
p<0,05 |
|
211* |
31 |
0,75 |
1,776 |
p<0,05 |
|
285 |
26 |
0,58 |
0,417 |
p>0,05 |
Примечание: «*» — особи, научившиеся выбирать подкрепляемый стимул.
Во втором эксперименте («выбор меньшего пятна») при тестировании в режиме с короткими МТИ 4 зверька из 9 (44,4%) продемонстрировали научение, им потребовалось 25—28 тестов (табл. 3). На уровне тенденции доля научившихся зверьков меньше, чем научившихся в первом эксперименте при том же режиме, но различия недостоверны.
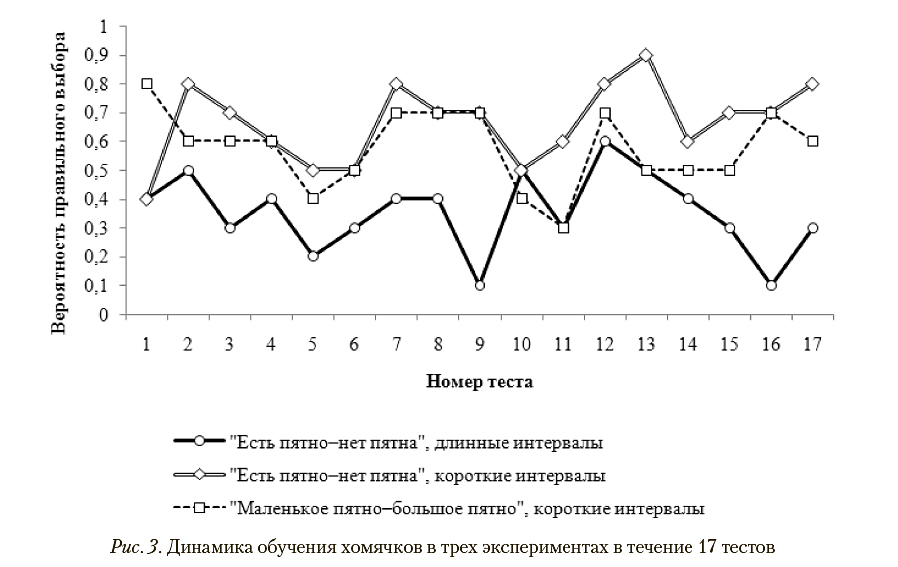
Мы сопоставили динамику обучения хомячков при одинаковом количестве тестов (17) во всех трех экспериментах (рис. 3).
Вероятность правильного выбора — доля выборов подкрепляемого стимула во всех «экзаменах» всех протестированных в этом тесте животных.
Видно, что вероятность правильного выбора при обучении с длинными МТИ постоянно ниже, чем в режиме обучения с короткими МТИ, а задачу «есть пятно—нет пятна» успешно решало большее или равное количество животных по сравнению с задачей «маленькое пятно—большое пятно». Таким образом, в течение первых 17 тестов при тестировании в режиме с короткими МТИ животные демонстрировали большую успешность обучения, чем в режиме с длинными МТИ.
Таблица 3
Доля выборов подкрепляемого визуального стимула в 25—28 «экзаменах» у джунгарских хомячков при тестировании с короткими МТИ
|
Номер животного |
Количество тестов |
Доля правильных выборов |
Значение биномиального критерия, Z |
Уровень значимости |
|
4 |
25 |
0,47 |
1,029 |
p>0,05 |
|
8* |
27 |
0,75 |
0,532 |
p>0,05 |
|
124* |
25 |
0,84 |
1,715 |
p<0,05 |
|
125* |
29 |
0,76 |
0 |
p>0,05 |
|
152* |
34 |
0,74 |
— 0,686 |
p>0,05 |
|
153* |
25 |
0,92 |
2,833 |
p<0,05 |
|
184* |
25 |
0,88 |
2,155 |
p<0,05 |
|
211* |
31 |
0,75 |
1,776 |
p<0,05 |
|
285 |
26 |
0,58 |
0,417 |
p>0,05 |
Примечание: «*» — особи, научившиеся выбирать подкрепляемый стимул.
В контрольном эксперименте ни одно из 9 животных не продемонстрировало достоверного предпочтения одного направления другому (биномиальный критерий, p>0,05).
Таблица 4
Доля выборов контейнера слева/справа в 10 «экзаменах» у джунгарских хомячков
|
Номер животного |
Доля выборов |
|
|
Контейнера слева |
Контейнера справа |
|
|
4 |
6 |
4 |
|
8 |
5 |
5 |
|
124 |
4 |
6 |
|
125 |
6 |
4 |
|
152 |
5 |
5 |
|
153 |
6 |
4 |
|
184 |
3 |
7 |
|
211 |
7 |
3 |
|
285 |
5 |
5 |
Обсуждение и заключение
В большинстве исследований, выполненных в «когнитивной» парадигме, более эффективным признается использование длительных интервалов (обзор см: Smolen et al., 2016), но накапливаются и противоположные свидетельства, связанные в основном с использованием разных генетических линий грызунов. Так, для мышей линии Fmr1 KO в задаче узнавания нового объекта эффективным оказалось обучение с длительными МТИ, а мыши дикого типа успешно обучались как в одном длинном тесте (t=5 мин), так и в трех коротких (t=100 с), разделенных длинными 1 час МТИ (Seese et al., 2014). В той же экспериментальной схеме у крыс линии Wistar эффективность обучения с короткими МТИ (10 мин) не уступала эффективности обучения с длительными МТИ ( 24 часа), хотя время, требуемое для консолидации памяти, различалось (Bello-Medina et al., 2013). В работе О.В. Перепелкиной и соавторов (Perepelkina et al., 2015) мыши линии ЭКС, селектированные на решение экстраполяционных задач, достигали высоких результатов при обучении с относительно МТИ (6 тестов в течение одного экспериментального дня). Как уже отмечалось, разные исследователи понимают под «длинными» и «короткими» МТИ отрезки времени, различные не только для разных типов и классов животных (насекомые, моллюски, млекопитающие), но и для одного отряда: как видно из приведенных выше исследований, в работе с грызунами «длинными» могут считаться интервалы от 1 до 24 часов.
В наших экспериментах с джунгарскими хомячками в режиме обучения с длинными МТИ (24 часа) способность связывать подкрепляемый визуальный стимул (наличие/ отсутствие черного пятна на белом фоне) с вознаграждением проявил один зверек из 10. Ранее при том же режиме тестирования с предъявлением более сложной задачи (дифференциация хорошо различаемых множеств геометрических фигурок: 5 и 10) были получены сходные результаты: научились 2 из 30 хомячков (Роговая и др., 2016). При режиме обучения с короткими МТИ способность к научению той же задаче («есть пятно/нет пятна») проявили 7 из 9 животных. При небольшом усложнении задачи (выбор меньшего по размеру пятна) количество научившихся зверьков (на уровне тенденции) было меньше: 4 из 9. Интересно отметить, что все особи, успешно научившиеся в эксперименте «выбор
меньшего пятна», ранее успешно научились в эксперименте «есть пятно/нет пятна» в режиме обучения с короткими МТИ, а один хомячок (№ 124) успешно связал визуальный стимул с подкреплением во всех трех сериях тестов. Эти, хотя и ограниченные, данные позволяют предположить, что распределение когнитивных способностей в популяциях джунгарских хомячков неравномерно, и возможно даже говорить о когнитивной специализации как одной из составляющих индивидуальной изменчивости (Резникова, 2011). Этот вопрос требует дальнейших исследований. В целом же, судя по нашим данным, когнитивные компетенции джунгарского хомячка в детализации визуальных стимулов («выбор меньшего пятна» ) и тем более их количественной оценке (дифференциация даже хорошо различаемых множеств) весьма ограничены. Это существенно отличается от результатов, полученных нами ранее на полевых мышах (Vorobyeva et al., 2013), которых по проявлению способностей к детализации и количественной оценке визуальных стимулов можно поместить на «другой конец шкалы». При оперировании визуальными стимулами у хомячков оказалась «короткая память». По-видимому, за редким исключением, они могут сформировать ассоциацию между визуальным стимулом и подкреплением только в режиме с короткими МТИ.
Вполне возможно, что когнитивные возможности хомячков в большей степени раскроются при предъявлении им задач с ольфакторными стимулами, как это сделано в работах А.В. Сурова (2006) на разных видах хомячков и в исследовании по связыванию ольфакторного стимула с подкреплением на лабораторных мышах (Kermen et al., 2010).
В целом можно сказать, что в экспериментальных исследованиях влияния режимов обучения на формирование ассоциативных связей между визуальными стимулами и подкреплением джунгарские хомячки могут служить своеобразной точкой отсчета, как животные, достигающие успеха только при режиме обучения с короткими МТИ.
Финансирование
Исследования поддержаны грантами РФФИ (№17-04-00702) и Программ ФНИ государственных академий наук на 2013—2020 гг., № VI.51.1.10. (АААА-А16-116121410120-0), №0-109-2018-0074.