Введение
Важное место в изучении проблемы соотношения психического и физиологического отводится решению вопроса сопряжения процессов формирования ощущения и реализации собственно эффекторной реакции, например, зрительно-моторной [Барабанщиков, 2009; Иваницкий, 2010; Иваницкий, 2017; Костандов, 2014; Block Ned. Consciousness, 2007; Clark, 2000; Woods, 2015]. В фундаментальных работах ряда авторов разрабатываются теории формирования ощущения, восприятия и осознания, построенные на так называемом принципе «кольца» А.М. Иваницкого [Иваницкий, 1996; Иваницкий, 2010], «динамического ядра» Дж. Эделмена [Edelman, 2000], а также на результатах исследований В.Я. Сергина [Сергин, 2016; Сергин, 2004], Ф. Крик [Crick Francis. The, 1994] и др. По данным А.М. Иваницкого [Иваницкий, 1996], ощущение формируется после нескольких возвратных циклов возбуждения, замыкающихся в первичной зрительной коре, с величиной кванта, равного 100—150 мс. Время повторного входа сигналов в модели Дж. Эделмена [Edelman, 2000] оценивается величиной до 300 мс, а латентность позднего визуального компонента на ЭЭГ, отражающего эффект перцепции, достигает 420 мс [Aalderen-Smeets, 2006; 30. Jeffreys D, 1986; Railo Henry, Koivisto, 2009]. Между тем известно, что время простой зрительно-моторной реакции (stimulus reaction time, sRT) колеблется в пределах 160—200 мс в зависимости от методического исполнения эксперимента [Соболев, 2019; Woods, 2015]. Причину возникновения такого рода временного разрыва (более 200 мс) позволяет объяснить гипотеза существования механизма разобщения процесса реализации моторного ответа при выполнении простой зрительно-моторной реакции, с одной стороны, и собственно процесса формирования осознанного светового ощущения — с другой. В этой связи представляется важным решение принципиального вопроса: необходимо ли осознание ощущения для запуска и выполнения ПЗМР и каковы механизмы ее реализации? Первые достоверные инструментальные исследования, проведенные в этом направлении, были выполнены Элизабет Ферер и Давидом Раабом [Fehrer, 1962]. Авторы установили, что время реакции на тестовый световой стимул в условиях обратной маскировки метаконтрастом соответствовало времени реакции в условиях без использования световой маскировки («Fehrer—Raab effect»); этот эффект сохранялся при изменении межстимульного интервала в диапазоне 0—75 мс. За верхними пределами этого интервала дополнительное раздражение, вызывающее возбуждение новых нервных элементов, уже не может помешать сенсорному осознанию первого, тестового, стимула [Бахманн, 1989; Костандов, 2014; Bachmann, 2009].
В классической работе E. Fehrer, D. Raab [Fehrer, 1962], так же как и в более поздних исследованиях разных авторов, использовали модель так называемой метаконтрастной световой маскировки [Bachmann, 2009; Bruchmann, 2010; Francis, 2000]. Такой способ маскировки имеет ряд достоинств, но и определенные недостатки, связанные, в частности, с ее черно-белым исполнением и в той или иной степени сложной конфигурацией предъявляемого зрительного стимула. Таких недостатков в значительной степени можно избежать в случае использования модели двухцветной ОСМ вспышкой света, когда цвет тестового и маскирующего стимулов различаются.
При изучении «Fehrer—Raab effect» [Fehrer, 1962] отдельный интерес представляет вопрос об особенностях его реализации при использовании сверхкоротких (десятки мкс) тестовых стимулов слабой интенсивности, а также вопрос о выборе для регистрации наиболее информативного показателя в составе простой зрительно-моторной реакции и наиболее точного метода его измерения. В последнем случае, с нашей точки зрения, предпочтение следует отдать параметру «время обнаружения стимула — stimulus detection time, sDT» [Woods, 2015], а для его измерения использовать электромиографический метод.
Наконец, представляется важным определение времени, необходимого для процесса консолидации следов тестового стимула в условиях ОСМ при формировании осознанного ощущения [Костандов, 2014]. На практике этот период вычисляется как время от начала предъявления тестового стимула до момента запуска маскера (stimulus onset asynchrony, SOA). В парадигме ОСМ после окончания этого периода повторная стимуляция маскером уже не будет оказывать существенного влияния на процесс осознания, а эффект ОСМ исчезает. В работе Э. Ферер и Д. Рааб [Fehrer, 1962] этот параметр ОСМ составил 75 мс, но не исключено, что в других условиях последний может кратно отличаться.
Целью работы явилось определение методом регистрации электромиограммы значения времени обнаружения хроматически разнородных сверхкоротких (0,1 мс) стимулов (stimulus detection time, sDT) в составе простой зрительно-моторной реакции, реализуемой в условиях блокады процесса формирования ощущения в парадигме полной обратной световой маскировки двухцветными стимулами, а также проведение анализа условий и закономерностей проявления маскировки.
Методика
Характеристика контингента испытуемых. Все исследования при соблюдении общепринятых этических норм были проведены с участием 14 девушек-студенток в осенний период 2017—2018 учебного года. Возраст испытуемых составлял 18,3±0,2 года.
Процедура и оборудование. В работе использовался классический алгоритм исследования эффектов ОСМ [Бахманн, 1989]. В качестве раздражителей предъявлялась пара светоимпульсов, состоящая из тестового стимула (1-й импульс — ТС) и маскера (2-й импульс — МС); предъявление осуществлялось последовательно с регулируемым интервалом времени (от начала ТС до начала МС, асинхрония включения стимулов, stimulus onset asynchrony, SOA).
В эксперименте № 1 (световая маскировка не использовалась) с помощью регистрации ЭМГ-ответа измеряли значение наиболее информативного параметра простой зрительно-моторной реакции (ПЗМР) — время обнаружения стимула (sDT) в ответ на изолированное предъявление красного (30 измерений), зеленого (30), а также случайным образом чередующихся красного (30) или зеленого (30) световых стимулов («красный/зеленый»). Во всех случаях длительность светоимпульсов составляла 0,1 мс.
В эксперименте № 2 определяли параметры стимуляции, при которых развивался максимально возможный (100%) эффект полной ОСМ тестового стимула последующей вспышкой синего света (маскера). В случае предъявления ТС и МС испытуемые должны были твердо идентифицировать факт проявления (ДА) или отсутствия (НЕТ) полной ОСМ. Предполагались только два ответа (ДА или НЕТ) на два вопроса: а) видят ли испытуемые одну световую вспышку (синий цвет МС, маскирующего ТС) в случае полной маскировки (ответ — ДА) или б) две раздельные цветоразличимые вспышки света (тестовый стимул и стимул маскер), т. е. эффект полной маскировки отсутствует (ответ — НЕТ). Уточнялось, что ответ должен быть однозначным (ДА или НЕТ), а в случае неопределенности цветоощущения испытуемым предлагалось отвечать «НЕТ». Маскировка считалась полной (100%), если испытуемый во всех пробах-тестах (10 попыток) однозначно отвечал «ДА». В случае отсутствия полной маскировки или сомнения в этом испытуемый отвечал «НЕТ» (эффективность ОСМ меньше 100%). В ходе опыта первоначально устанавливали равную длительность ТС и МС на уровне 0,1 мс (т. е. ТС и МС в шкале времени накладывались друг на друга, а SOA, следовательно, равнялась нулю). При таких параметрах эффект маскировки еще не проявлялся, а испытуемые всегда идентифицировали две цветоразличимые вспышки света. Затем длительность стимула-маскера (синего) увеличивали до 45 мс при ТС = 0,1 мс и SOA = 0, что вызывало эффект полной (100%) обратной маскировки. В дальнейшем при неизменной длительности ТС и МС (соответственно, 0,1 мс и 45 мс) SOA-интервал последовательно удлиняли от исходного уровня 0 мс до 5, 10, 15, 20, 25, 30, 35 или 40 мс (варианты теста). Количество предоставляемых попыток в каждом тесте-варианте при оценке факта «наличие маскировка/отсутствие маскировки» составляло 10.
В эксперименте № 3 через 30 мин после процедуры определения показателя степени полной ОСМ измеряли время обнаружения светового стимула при каждом значении SOA-интервала (0, 5, 10, 15, 20, 25, 30, 35 и 40 мс) и неизменных параметрах ТС = 0,1 мс и МС = 45 мс. Число измерений sDT при каждом значении SOA составляло 30 с паузами, изменяющимися в случайном порядке в диапазоне от 4 до 7 с. Описанный в экспериментах № 2 и № 3 цикл выполняли трижды для световых пар «ТС—МС» : «красный—синий», «зе- леный—синий» и «(красный/зеленый) — синий».
В блоке светостимуляции экспериментальной установки SOA-интервал между тестовым стимулом и стимулом-маскером мог быть выбран в диапазоне от нуля (1-й и 2-й импульсы накладывались друг на друга) до 40 мс. В качестве источника света использовали RGB-светодиод (Ultra brightness BL-L515RGBC), что позволило инициировать вспышку красного (Ultra Orange 630 nm), зеленого (Ultra Pure Green 525 nm) или синего (Blue 430 nm) света. В данном типе светодиода красный, зеленый и синий излучатели располагаются в углах равнобедренного треугольника со стороной не более 1 мм. С учетом достаточно большого расстояния сетчатки глаза до излучателя (40 см) и рассеивающей линзе (угол рассеивания 20°) стимулы ТС и МС предъявлялись в одну и ту же, хотя и обширную, область сетчатки. Значение силы света для каждого светового излучателя (красного, зеленого и синего) являлось постоянной величиной и устанавливалось на приемлемом для глаза уровне 2000 mcd (luminous intensity).
В блоке ЭМГ-канала с помощью биоусилителя (на базе INA118) и электронного самописца (S-Recoder-L, Россия) регистрировалась электромиограмма, запись которой в дальнейшем использовалась для вычисления параметра ПЗМР — «времени обнаружения стимула (sDT»). Численно значение параметра sDT соответствовало отрезку времени (рис. 1) от момента предъявления сенсорного стимула до момента появления первых ЭМГ- потенциалов (ЭМГ-ответ). Для регистрации ЭМГ использовали ЭМГ-электроды диаметром 10 мм, которые крепились на коже (4 см друг от друга) над мышцами внешней поверхности предплечья, участвующих в сгибании пальцев кисти.
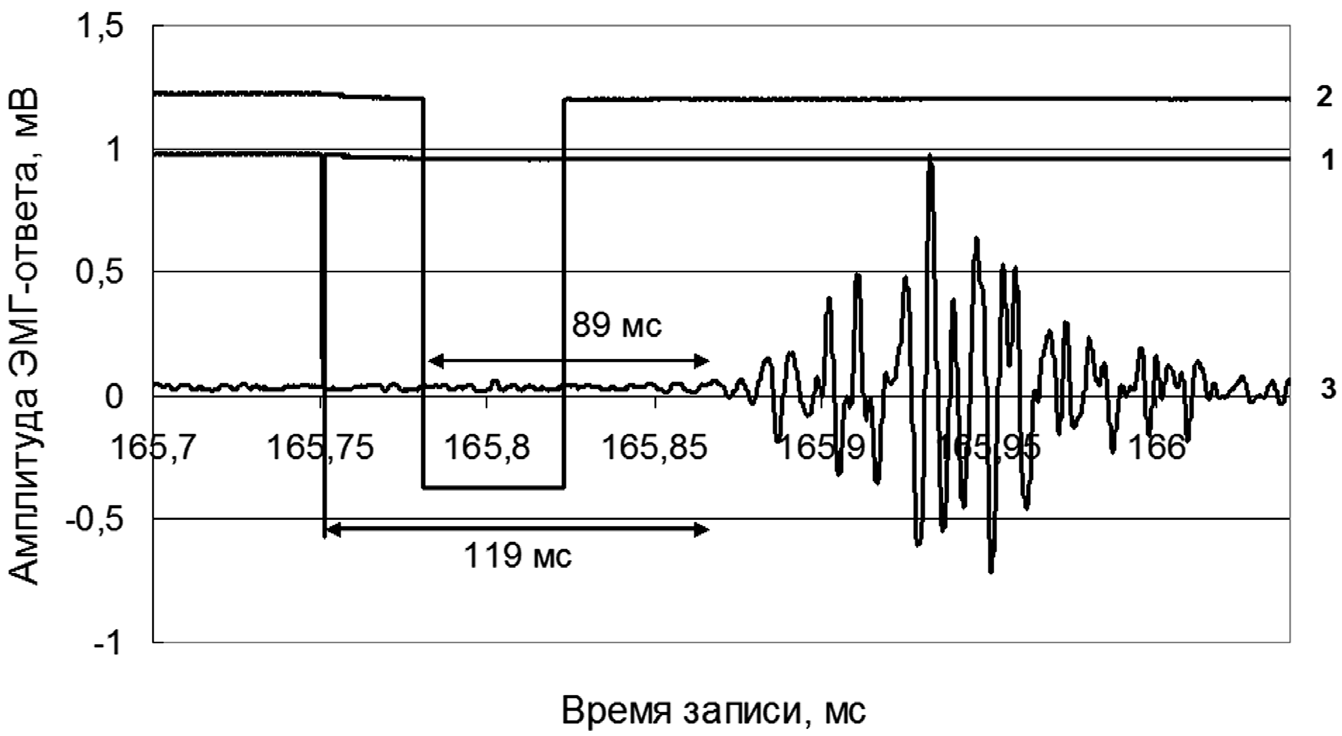
Рис. 1. К определению параметра «время обнаружения стимула — sDT» при выполнении простой зрительно-моторной реакции методом регистрации ЭМГ-ответа в парадигме обратной световой маскировки: 1 — тестовый стимул длительностью 0,1 мс; 2 — стимул-маскер длительностью 45 мс; 3 — электромиограмма (ЭМГ-ответ); SOA-интервал (по переднему фронту световых импульсов) равен 30 мс; вычисленное значение параметра sDT составляет 119 мс; условие измерения (состояние) — полная обратная световая маскировка вспышкой света
В ходе опыта в затемненной комнате правая рука испытуемого с наложенными ЭМГ- электродами в положении «сидя» свободно располагалась вдоль тела, что обеспечивало низкий уровень исходного ЭМГ-тонуса, а реакция на сенсорный стимул осуществлялась путем максимально быстрого встречного сведения большого и указательного пальцев кисти.
Статистический анализ. При статистической обработке данных использовали пакеты прикладных программ Excel и Statistica 7.0. После определения характера распределения данных в сводных вариационных рядах (тест Шапиро—Уилка) рассчитывали основные показатели описательной статистики. Статистическую значимость различий между двумя средними арифметическими величинами определяли с помощью двухвыборочного t-теста Стьюдента для выборок с различными дисперсиями при заданном уровне значимости p < 0,05. При оценке различий между двумя множествами использовали двухвыборочный F-тест для дисперсий. Для характеристики вариабельности исследуемых показателей рассчитывали величины дисперсии выборок и коэффициенты вариации. Во всех случаях сравнение анализируемых показателей и оценку статистической значимости различий проводили на основании проверки нулевой и альтернативной гипотез.
Результаты
Эксперимент № 1. Анализ показал (табл.), что время обнаружения светового стимула (sDT) при выполнении ПЗМР составило для вспышки красного света 141±1,1 мс, зеленого — 142±1,2 мс и при их чередовании 143±1,5 мс, т. е. статистически значимо не различалось (p > 0,05). Отсутствовали различия и со стороны коэффициентов вариации (3,5—4,1%) и величины дисперсий (p > 0,05). Характер распределения данных в вариационных рядах описывался нормальным законом.
Эксперимент № 2. На следующем этапе исследований были найдены параметры стимуляции (длительность стимула-маскера и значение SOA-интервала), при которых проявлялась световая маскировка тестовых световых раздражителей (красного или зеленого) вспышкой синего света стимула-маскера (табл.). Анализ показал, что эффект полной ОСМ («Вижу только синюю вспышку света») легко достижим, начиная с минимального в наших исследованиях SOA-интервала 0 мс при длительности стимула-маскера 45 мс. Зона полной ОСМ простиралась в диапазоне SOA от 0 до 25 мс, сменяясь в дальнейшем зоной ее угасания (30—40 мс). Например, при алгоритме «Green—Blue» зона угасания начиналась при достижении SOA значения 30 мс, о чем свидетельствует уменьшение степени эффективности ОСМ до (65,9±7,6%). Во всех модификациях опыта значение точки начала угасания ОСМ стимула было одинаковым и соответствовало SOA-интервалу 30 мс. Следовательно, длительность критического периода, необходимого для завершения начального этапа процесса осознания хроматических стимулов в парадигме двухцветной ОСМ, во-первых, для всей группы испытуемых (100%) составила 25 мс и, во-вторых, не зависела от цвета тестового стимула. При SOA, равной 40 мс, эффект ОСМ во всех случаях полностью исчезал (0%).
Эксперимент № 3. После нахождения параметров и условий развития ОСМ было измерено время обнаружения стимула как составной части ПЗМР. Результаты экспериментов показали, что величина показателя sDT не зависела от параметров стимуляции. Например, в случае реакции на изолированный («одиночный») тестовый стимул красного света значение sDT составило 141±1,1 мс, а при использовании стимула-маскера минимальной длительности (при комбинации «0,1—0,1—0 мс», табл.), когда эффект полной маскировки еще не проявлялся, время реакции статистически значимо (p = 0,83) не изменилось (140±1,2) мс.
В условиях развития обратной световой маскировки разной степени выраженности значение вычисленного показателя «время обнаружения стимула» оставалось на неизменном уровне. Так, при полной (100%) ОСМ и длительности SOA-интервала (0—40 мс) величина sDT для модели «Red—Blue» колебалась в пределах 140—142 мс, а в случае «Green— Blue» — в диапазоне 141— 45 мс, т. е. статистически значимо не различалась (p < 0,05). Аналогичная закономерность имела место и в условиях угасания обратной световой маскировки вплоть до ее полного исчезновения (табл.). Сделанный вывод подтверждается также результатами дисперсионного анализа (двухвыборочный F-тест для дисперсий): во всех случаях вариационные ряды характеризовались статистически равными дисперсиями.
Таким образом, анализ результатов эксперимента свидетельствует, что время обнаружения стимула при выполнении ПЗМР при обратной маскировке вспышкой света не зависит ни от эффективности ОСМ, ни от цвета тестового стимула, ни от величины асинхронии включения стимулов, ни от длительности стимула-маскера, а инициация сенсомоторной реакции не нуждается в осознании светового ощущения.
Обсуждение результатов
Анализ результатов исследований показал, что модель двухцветной ОСМ может быть использована для анализа закономерностей сопряжения процессов формирования ощущения и реализации сенсомоторных реакций. Следует отметить, что эффект ОСМ преимущественно изучался на ее метаконтрастной модели [11, 17, 21, 22, 23, 25]. По нашим данным, эффект полной (100%) двухцветной ОСМ надежно реализуется при следующих ее параметрах: длительность тестового стимула (красный и зеленый цвета) 0,1 мс, стимула-маскера (синий) 45 мс и SOA 0—25 мс; в диапазоне значений SOA от 25 до 35 мс эффективность ОСМ, т. е. число случаев полной маскировки, снижается до 14%, а при SOA 40 мс эффект полной маскировки полностью исчезает.
Таблица
Значение параметра «время обнаружения стимула — sDT» простой зрительно-моторной реакции (M±m) при разных значениях параметров стимуляции в условиях обратной световой маскировки двухцветной модели
|
Параметры стимуляции (ТС—МС— SOA), мс |
Условия эксперимента (цвет тестового стимула и стимула-маскера) |
|||||
|
«Red—Blue» |
«Green—Blue» |
«Red/Green—Blue» |
||||
|
Среднее и ошибка |
Показатель степени полной маскировки, (%) |
Среднее и ошибка |
Показатель степени полной маскировки, (%) |
Среднее и ошибка |
Показатель степени полной маскировки, % |
|
|
0,1—нет—нет |
141±1,1 |
- |
142±1,2 |
- |
143±1,5 |
- |
|
0,1—0,1—0 |
140±1,2 |
0 |
140±1,3 |
0 |
142±1,4 |
0 |
|
0,1—45—0 |
142±1,5 |
100 |
141±1,2 |
100 |
144±1,6 |
100 |
|
0,1—45—5 |
140±1,4 |
100 |
142±1,5 |
100 |
144±1,6 |
100 |
|
0,1—45—10 |
141±1,2 |
100 |
143±1,4 |
100 |
143±1,6 |
100 |
|
0,1—45—15 |
142±1,3 |
100 |
143±1,3 |
100 |
145±1,4 |
100 |
|
0,1—45—20 |
140±1,6 |
100 |
144±1,4 |
100 |
143±1,5 |
100 |
|
0,1—45—25 |
141±1,2 |
100 |
142±1,5 |
100 |
145±1,6 |
100 |
|
0,1—45—30 |
142±1,5 |
68,6±4,0 |
145±1,6 |
65,9±7,6 |
144±1,7 |
77,5±4,5 |
|
0,1—45—35 |
140±1,4 |
12,4±3,8 |
144±1,4 |
14,1±5,3 |
145±1,7 |
20,1±5,8 |
|
0,1—45—40 |
142±1,6 |
0 |
143±1,6 |
0 |
144±1,8 |
0 |
Примечание: ТС — тестовый стимул (Red — красный, Green — зеленый); МС — маскер-стимул (Blue, синий); SOA — асинхрония включения стимулов (stimulus onset asynchrony).
Важным параметром ОСМ является «критическое время асинхронии включения стимулов, SOA ». Этот показатель отражает длительность периода абсолютной уязвимости процесса формирования осознанного ощущения (в нашем случае ощущения красного и зеленого цвета). В пределах SOA^ следующий за тестовым стимулом маскер другого цвета (синего) блокирует указанный перцептивный процесс. В наших исследованиях SOAk|j[it составил 25 мс, хотя для некоторой части испытуемых, например, при модели «Red—Blue» он растягивался до 30 мс (68,6±4,0 %) и даже до 35 мс (12,4±3,8%). В работах с применением черно-белой метаконтрастной маскировки абсолютное значение указанного параметра полной ОСМ определяется в диапазоне от 75 мс [Fehrer, 1962] до 100 мс и более [2, 6]. Существенные (кратные) различия по сравнению с нашими результатами (25 мс) связаны, как мы полагаем, с использованием разнотипных моделей ОСМ и обусловлены, по крайней мере, двумя обстоятельствами. Во-первых, при метаконтрастной маскировке фотостимуляции подвергаются топографически разобщенные участки сетчатки, тогда как в наших исследованиях ТС и МС предъявлялись в одну и ту же ее область. Во-вторых, принципиально различались и хроматические параметры моделей. Так, в случае монохромной метаконтрастной маскировки стимуляции подвергаются преимущественно одни и те же пулы хроматически однотипных фоторецепторов, еще не способных в течение определенного времени после реакции на тестовый стимул к повторному возбуждению. В результате SOA^ удлиняется минимум на значение длительности периода рефрактерности фоторецепторов. При двухцветной модели ОСМ импульсами света с несовпадающей длиной волны, как в нашем случае, последовательно стимулируются преимущественно хроматически разноименные пулы фоторецепторов, что исключает влияние данного фактора (абсолютной рефрактерности) на общее время SOA . Возможны и другие механизмы удлинения SOA^ включения стимулов в условиях монохромной ОСМ.
Таким образом, определенное в наших исследованиях значение SOA^ в модели двухцветной ОСМ указывает на относительно короткую (25 мс) продолжительность фазы абсолютной уязвимости процесса формирования осознанного ощущения (предсознательной компоненты ощущения).
Что же касается механизмов процесса прерывания цепочки формирования ощущения при обратной маскировке, то данная проблема имеет самостоятельный интерес и требует отдельных исследований фундаментального порядка. Тем не менее, следует отметить, что на этот счет существует более десятка различных теорий [10, 16, 22, 27]. В настоящее время список теорий и гипотез маскировки еще более расширился за счет экспериментального использования новейших методологических и методических подходов, в том числе транскраниальной магнитной стимуляции и других высокоинформативных методик [26, 27, 28], включая построение математических моделей маскировки [Francis, 2000].
Центральной задачей нашей работы явилось измерение значения времени обнаружения хроматически разнородных сверхкоротких (0,1 мс) стимулов (sDT) в условиях блокады процесса формирования сенсорного ощущения в парадигме полной ОСМ. Результаты показали, что параметр sDT не зависит от эффективности ОСМ (от 0 до 100%), цвета тестового стимула («Red» или «Green», 0,1 мс), величины асинхронии в диапазоне (0—40 мс), длительности стимула-маскера («Blue», от 0,1 до 45 мс), а также не нуждается в формировании осознанного светового ощущения. Представляют интерес результаты сравнительного анализа времени обнаружения стимула в условиях без использования ОСМ и при ее полном проявлении. По нашим данным, абсолютное значение показателя sDT при полной ОСМ (например, при алгоритме «Red—Blue») составляло 140—142 мс (табл.), а при ее отсутствии 140 мс, т. е. не изменялось. Следовательно, «Fehrer—Raab-effect» проявляется не только в парадигме черно-белой метаконтрастной ОСМ, но в полной мере сохраняется и при двухцветной маскировке для сверхкоротких хроматически разнородных тестовых стимулов.
Феномен независимости запуска и выполнения ПЗМР от цвета стимулов и процесса формирования осознанного светового ощущения предполагает существование двух автономных путей ее реализации. Во-первых, механизм ОСМ при ее двухцветной модели не связан с периферическим звеном зрительного анализатора. Такое предположение имеет право на существование, если доказать, что информационный поток, рожденный тестовым стимулом, успевает покинуть рецепторно-сетчатый аппарат зрительного анализатора еще до момента предъявления испытуемому стимула-маскера. Разумеется, что наилучшим вариантом было бы проведение прямой регистрации потенциалов действия, например, в зрительном нерве. Однако существуют и другого рода возможности. Первая из них основана на результатах измерения латентности вызванных зрительных потенциалов в первичной зрительной коре головного мозга при фотостимуляции. Известно, что латентность первых волн на электроэнцефалограмме (P1 и N1) нередко находится в пределах 30—50 мс. В известной схеме «кольца А.М. Иваницкого» [Иваницкий, 1996] появление вызванного потенциала в первичной зрительной коре относится к 30 мс. Подчеркнем, что в наших исследованиях в эксперименте № 3, наряду с другими вариантами, время между предъявлением ТС и МС устанавливалось на уровне 30 мс, т. е. на временном отрезке, достаточном для физического поступления информационного потока от тестового стимула с периферического звена анализатора до его центральных отделов. Понятно, что в таких условиях стимул-маскер, предъявляемый, в частности, через 30 мс после ТС, не может инициировать запуск механизма обратной маскировки, которую при использованных параметрах ОСМ мы всегда наблюдали. Такого рода данные не подтверждают периферическую природу ОСМ. В качестве второго, хотя и косвенного доказательства тезиса, опровергающего периферическую природу механизма ОСМ, можно привести одну из записей ЭМГ-ответа, произведенной в условиях полной обратной маскировки (ТС = 0,1 мс, МС = 45 мс и SOA = 30 мс) при вычисленном значении sDT = 119 мс (рис. 1). Результаты анализа показывают, что если бы запуск ПЗМР осуществлялся в ответ на предъявление МС, то гипотетически латентный период реакции (sDT) составил бы 89 мс, однако таких показателей времени получено не было: при всех вариантах опытов эмпирическое значение sDT колебалось в пределах 118—150 мс при средней величине 141 мс. Кроме того, согласно литературным данным [9, 29], корректно измеренное время ПЗМР всегда превышает 120 мс. Следовательно, ПЗМР в наших экспериментах в условиях полной ОСМ инициировалась не маскером, а тестовым стимулом, что возможно лишь при условии опережающего «выхода» потока нервных импульсов из рецепторно-сетчатого аппарата. В связи с этим можно полагать, что структурно-функциональной областью, на которую направлено действие механизма ОСМ использованной двухцветной модели, является не периферическое звено зрительного анализатора, а его центральные отделы.
Во-вторых, факт запуска и выполнения ПЗМР вне зависимости от формирования соответствующего ощущения свидетельствует о том, что механизм реализация ПЗМР и механизм формирования ощущения разобщены как во времени проявления, так, скорее всего, и в отношении структур мозга, на которых их действие распространяется. Можно полагать, что, с одной стороны, существует укороченный отдельный скоростной тракт для реализации ПЗМР, используемый для простого обнаружения светового сигнала как такового без идентификации его характеристик (цвета и интенсивности) и необходимости осознания, а с другой стороны — путь, предполагающий, в конечном итоге, формирование осознанного светового ощущения. Оба механизма в нативных условиях функционируют параллельно, но с разной латентностью, а в условиях ОСМ скоростной механизм простого обнаружения сигнала функционирует сугубо изолированно, не предполагая непременного формирования осознанного ощущения.
Бессознательная реакция на тестовый стимул предполагает участие отдельной функциональной системы, обеспечивающей реализацию всего комплекса зрительно-моторного ответа. Мы полагаем, что такая функциональная система, вернее ее шаблон, хранится в сенсорной памяти и сформирована ранее на основании предшествующего индивидуального эмпирического опыта. В ходе словесной установки экспериментатора в виде описания алгоритма поведенческой реакции испытуемого такого рода функциональная система корректируется в соответствии с новыми условиями ее выполнения, несколько раз (5—10) апробируется в процессе начального цикла обучения, закрепляясь в оперативной памяти, переходит в режим активного ожидания, а в случае предъявления светового стимула запускается по принципу триггера. Однако обсуждаемый аспект проблемы нейрофункциональной организации «Ферер—Рааб-феномена» в условиях ОСМ является отдельной задачей и требует фундаментальных исследований нейрофизиологического характера [4, 6, 10, 14].
Выводы
1. Модель обратной маскировки вспышкой света в ее двухцветной модификации (ОСМ) применима при исследовании механизмов сопряжения процессов формирования ощущения и реализации сенсомоторных реакций. Эффект полной (100%) двухцветной ОСМ надежно реализуется при следующих значениях ее параметров: длительность тестового стимула (красный и зеленый цвета) — 0,1 мс, длительность стимула-маскера (синий) — 45 мс, диапазон асинхронии включения стимулов (SOA) 0 — 25 мс. В диапазоне значений SOA от 25 до 35 мс степень проявления полной ОСМ снижается до 12—14%, а при SOA, равном 40 мс, полностью исчезает. Вычисленное критическое значение SOA (25 мс) в модели двухцветной ОСМ указывает на относительно короткую длительность фазы абсолютной уязвимости процесса формирования осознанного ощущения в центральных структурах зрительного анализатора на стадии консолидации следов светового стимула.
2. Время обнаружения стимула (sDT) в составе простой зрительно-моторной реакции в парадигме модели двухцветной обратной световой маскировки не зависит от ее эффективности (от 0 до 100%), цвета тестового стимула («Red» или «Green», 0,1 мс), величины асинхронии в диапазоне (0—40 мс), длительности стимула-маскера («Blue», от 0,1 до 45 мс), а также не нуждается в формировании осознанного светового ощущения.
3. Простая зрительно-моторная реакция в парадигме двухцветной обратной световой маскировки всегда инициируется первым, тестовым стимулом (0,1 мс), а ее запуск осуществляется вне связи с формированием осознанного светового ощущения. Сравнительный анализ значений sDT в условиях полной (100%) ОСМ и при ее отсутствии позволяет предполагать, что механизм обратной световой маскировки при ее двухцветной модели не связан с периферическим рецепторным звеном зрительного анализатора, а реализуется, следовательно, в его центральных отделах.