Введение
Фундаментальные проблемы психологии восприятия по-прежнему остаются в сфере внимания психологии, психофизиологии, нейрофизиологии, неврологии, психиатрии и педагогики [Барабанщиков, 2019; Иваницкий, 2010; Костандов, 2014; Масленникова, 2020; Шиффман, 2003]. Однако, несмотря на многолетнюю и плодотворную историю изучения, многие аспекты теории восприятия остаются недостаточно ясными, в частности, это относится к проблеме измерения времени формирования осознанного ощущения и отдельных его компонентов [Иваницкий, 1996; Костандов, 2014; Шиффман, 2003; Rutikua, 2016]. Следует отметить, что определение данного показателя осложняется самой природой исследуемого феномена и, прежде всего, его субъективной оценкой со стороны индивида. Перед исследователем всегда возникает вопрос о степени синхронности возникновения осознанного ощущения и «подачей знака» испытуемым о подобном событии. Кроме того, феномен ощущения проявляется не мгновенно, а занимает определенное трудноопределимое время. Задача еще больше усложняется, если возникает необходимость измерения латентности осознания кратковременных сенсорных стимулов, например, длительностью в несколько сотен мкс.
Среди перспективных методологических подходов при исследовании указанной проблемы восприятия можно выделить нейрофизиологические методы, основанные на определении латентности и топографии вызванных потенциалов, связанных с теми или иными событиями [Иваницкий, 1992; Railo, 2009; Sysoeva, 2007]. Однако и в этом случае остается неясным насколько точно, например, те или иные N- и P-волны, синхронизированы с моментом начала появления у испытуемого собственно осознанного ощущения. Необходимо также учитывать, что в случае длительности N- и P-волны, равной 100—150 мс [Бахманн, 1989], определение точного времени момента проявления ощущения затруднено. Ряд авторов считают, что латентность визуального компонента на ЭЭГ, отражающего момент осознания предъявленного стимула или перцепции в зависимости от использованного метода исследования, находится в диапазоне от 160 до 420 мс [Baars, 1993; Beckers; Fahrenfort, 2008; Railo, Koivisto M, 2011; Railo, 2009; Rutikua, 2016; Sysoeva, 2007]. Применение других современных методов, например таких, как функциональная магниторезонансная томография [Koivisto, 2011], ограничено их значительной инерционностью. Наиболее адекватные прямые микроэлектродные исследования в условиях insitu в силу понятных причин (за исключением медицинских показаний) с участием человека не проводятся.
Используемый в психофизиологии метод измерения латентного периода зрительно-моторной реакции (ЗМР) для определения времени формирования осознанного ощущения в классическом исполнении малопригоден, так как позволяет измерить лишь комплексное время реакции, в которой латентности формирования ощущения принадлежит лишь определенная, не поддающаяся точному вычленению доля. Имеющаяся возможность несколько сузить рамки диапазона путем исключения из времени полной ЗМР моторного компонента и точного измерения параметра «время обнаружения стимула (stimuli detection time») [Соболев, 2020] в целом задачу не решает.
В литературе, касающейся психо- и нейрофизиологических основ восприятия, можно найти немалое число логико-структурных схем, описывающих как в целом, так и в отдельных деталях процесс формирования осознанного ощущения [Иваницкий, 1996; Иваницкий, 2010; Нехорошкова, 2015; Сергин, 2016; Шиффман, 2003; Miller, 2001]. Их описание и анализ представляют особый интерес, однако требуют отдельного аналитического обзора. Тем не менее, в настоящее время не вызывает сомнения, что для проявления осознанного ощущения (психического феномена — ПФ) необходимо первоначальное формирование физиологического феномена (ФФ), который в дальнейшем в соответствующем виде транслируется в центральные структуры зрительного анализатора в качестве объекта консолидации следов светового стимула [Костандов, 2014]. ФФ на разных стадиях своего создания (от уровня рецепторно-сетчатого аппарата до корковых отделов головного мозга) характеризуется специфическим нейрофизиологическим паттерном. Значение всех этих областей заключается в постепенном выявлении и анализе различных аспектов светового стимула [Измайлов, 1989; Шиффман, 2003]. На конечной стадии формирование ФФ полностью завершается, после чего он включается в процессы организации собственно осознанного ощущения. К сожалению, характеристика терминальной морфофункциональной нейронной структуры, в которой завершается процесс создания ФФ, равно как и продолжительность этой фазы, достоверно не установлены.
Задача определения времени формирования физиологического феномена в цепочке реализации процесса осознанного светового ощущения в психологии и психофизиологии восприятия в контексте настоящего исследования, по-видимому, не ставилась. В связи с этим аппаратное измерение значения этого показателя методами психофизиологического анализа, с одной стороны, представляет интерес для изучения природы и механизма реализации предсознательной фазы ощущения, а с другой — упрощает задачу определения длительности периода формирования одного из фундаментальных проявлений психики —феномена осознанного ощущения.
В качестве экспериментальной модели для измерения времени формирования физиологического феномена, как мы предлагаем, может быть использована дифференцировочная зрительно-моторная реакция «Go/No-go» типа (ДЗМР), реализуемая в парадигме двухцветной обратной маскировки [Соболев В.И, 2020]. Суть предложенного подхода состоит в следующем. Априори считается, что при ДЗМР «Go/No-go» типа моторный ответ запускается лишь при условии предварительной осознанной идентификации испытуемым факта предъявления тестового стимула (альтернативная точка зрения может предполагать, что реализация ДЗМР осуществляется по другому принципу, а осознанное ощущение проявляется постфактум). После выполнения всех последующих нейрональных операций (сравнение с заданным в эксперименте эталоном, принятие решения о способе реакции и др.) происходит инициация моторной программы и собственно моторного ответа. Если каким-либо способом затруднить (ослабить интенсивность, помешать, затормозить, исказить) процесс формирования физиологического феномена, как предиктора осознанного ощущения, то латентный период целостной, интегральной, зрительно-моторной реакции должен автоматически удлиняться. В качестве такого рода тормозящего фактора, с нашей точки зрения, можно использовать эффект стимула-маскера той же модальности, но другого цвета, который должен предъявляться на разных этапах организации зрительно-моторной реакции, но обязательно после завершения периода так называемого «критического времени асинхронии включения стимулов» — АВСкрит (Stimulus Onset Asynchrony, SOA). Этот показатель в парадигме обратной маскировки отражает длительность периода абсолютной блокады (эффект прерывания) процесса последующего формирования осознанного ощущения [Бахманн, 1989; Костандов, 2014]. Согласно нашей рабочей гипотезе, предполагается, что в тот момент, когда создание физиологического феномена будет близко к завершению или же закончено, тормозной эффект стимула-маскера на скорость ДЗМР вначале существенно ослабнет, а затем полностью исчезнет. Определив для данного момента значение времени асинхронии включения стимулов, можно будет судить о длительности фазы формирования физиологического феномена (АВСФФ).
Целью работы явилось определение длительности фазы формирования физиологического феномена как предиктора осознанного светового ощущения на основании результатов измерения латентности зрительно-моторной реакции «Go/No-go» типа, выполняемой на фоне предъявления в разные периоды ее реализации «сенсорной помехи» в виде стимула-маскера.
Методика
Характеристика контингента испытуемых
Исследования проведены в осенне-зимний период с участием 12 студентов-психологов (девушки) в возрасте 18—19 лет (M = 18,3; SD = 0,62) Института педагогики, психологии и инклюзивного образования Крымского федерального университета имени В.И. Вернадского.
Процедура исследования и оборудование
В эксперименте № 1 (табл. 1) выявляли параметры стимуляции, при которых развивался максимально возможный (100%) эффект полной обратной световой маскировки (ОСМ). В качестве раздражителей использовали пару светоимпульсов, составленных из тестового стимула (1-й импульс — ТС, Синий), и стимула маскера (2-й импульс — маскер-стимул, МС, Зеленый). Стимулы предъявляли последовательно с регулируемым интервалом времени, численно равным периоду от начала ТС до начала МС (АВС-интервал, асинхрония включения стимулов — АВС, Stimulus Onset Asynchrony—SOA). В ходе опыта первоначально устанавливали равное значение длительности ТС и МС на уровне 0,1 мс, т. е. ТС и МС в шкале времени накладывались друг на друга, а АВС, следовательно, равнялась нулю. При таких значениях параметров эффект маскировки еще не проявлялся, а испытуемые всегда идентифицировали две цветоразличимые вспышки света: «Вижу две вспышки света — тестовый светоимпульс (Синий) и светоимпульс маскера (Зеленый)». Затем длительность стимула-маскера увеличивали до 30 мс при ТС = 0,1 мс и АВС = 0, что незамедлительно вызывало эффект полной (100%) обратной маскировки: «Вижу только одну вспышку зеленого цвета». В дальнейшем при неизменной длительности вспышек света ТС и МС (соответственно 0,1 мс и 30 мс) АВС-интервал последовательно удлиняли от исходного уровня 0 мс до 10, 15, 20, 25, 30 или 40 мс. Маскировка считалась полной (100%) в том случае, если испытуемый во всех пробах-тестах (10 попыток) безальтернативно отвечал «ДА», т.е. маскировка имела место, в чем он был полностью уверен. В случае отсутствия полной маскировки или сомнения в этом испытуемый отвечал «НЕТ». Например, если в восьми попытках испытуемый отвечал «ДА», а в двух — «НЕТ», то, следовательно, показатель степени маскировки (другими словами — эффективности) составлял 80%. В дальнейшем данные для всей группы объединяли и вычисляли параметры вариационной статистики.
В ходе эксперимента № 2 (проводился в течение двух дней) измеряли латентный период дифференцировочной зрительно-моторной реакции (Reaction Time, RT) при разном значении АВС-интервала. RT измеряли дважды: первый раз с использованием маскера (помехи) при разных значениях АВС и второй раз — без него, т. е. без маскировки. В работе использовали известную модель дифференцировочной реакции простого выбора так называемого «Go/No-go» – типа. Согласно алгоритму этой модели, при чередующемся предъявлении двух стимулов испытуемый на один из них (тестовый стимул — ТС, Синий) должен отвечать моторной реакцией, а на другой (тормозной дифференцировочный стимул, Красный) — его игнорировать. Процедура заключалась в следующем: длительность ТС и тормозного дифференцировочного стимула устанавливалась на уровне 0,1 мс, а стимула-маскера (МС, Зеленый) на уровне 30 мс. Латентный период зрительно-моторной реакции измерялся при значениях АВС от 40 мс до 50, 60, 70, 80, 90, 100, 150, 200 и 300 мс. Число измерений RT при каждом значении АВС составляло 50 при паузах между очередным предъявлением стимула 4—6 с.
В эксперименте № 3 испытуемые оценивали силу субъективного ощущения после предъявлении тестового стимула (ТС = 0,1 мс, Синий) на фоне действия стимула-маскера (МС = 30 мс, Зеленый) при трех значениях АВС-интервала — 40, 70 и 100 мс. В ходе одного цикла АВС-интервал сменялся последовательно в следующем порядке: 40, 70 и 100 мс (итого, 3 варианта в одном цикле). Число выполненных циклов составляло 10. Анкетирование проводилось после выполнения каждого из 10 циклов. Использовалась трехбалльная шкала: 1 бал — «ощущение тестового стимула слабо выражено»», 2 бала — «ощущение средней степени выраженности» и 3 балла — «ощущение ярко выражено».
В блоке светостимуляции экспериментальной установки была предусмотрена возможность программного выбора длительности световых вспышек в широком диапазоне значений: от 0,1 мс до 45 мс. АВС-интервал между тестовым стимулом и стимулом-маскером устанавливался в диапазоне от нуля (1-й и 2-й импульсы накладывались друг на друга) до 300 мс. Длительность тестового и дифференцировочного импульсов, а также значение АВС-интервалов и другие опции устанавливались аппаратно с помощью элементов цифровой логики. Экспериментальная установка включала следующие основные блоки: блок генерации случайных временных интервалов (в диапазоне от 4 до 6 с); блок задержки импульсов путем изменения с помощью электронных сопротивлений параметров задающей RC-цепочки (дискретно на: 0 — 10 — 15 — 20 — 30 — 40 — 50 — 60 — 70 — 80 — 90 — 100 — 150 — 200 — 300 мс); блоки формирования длительности импульсов; буферные устройства и RGB-светодиод.
Латентный период дифференцировочной зрительно-моторной реакции соответствовал интервалу времени от момента предъявления тестового стимула до момента фиксации кнопки «СТОП» секундомера, т. е. его остановки. Электронный секундомер, построенный на основе микроконтроллера ATmega 8515-16PU и LED-дисплея CA56-12SRWA, позволял измерять интервал времени с точностью до 0,25 мс/дел. Во время эксперимента пульт с кнопкой фиксировали между большим (сверху) и четырьмя (снизу) пальцами правой руки. Высокая эргономичность пульта позволяла в процессе выполнения моторной реакции при встречном движении пальцев (имитация хватательного рефлекса) существенно укорачивать время срабатывания кнопки «СТОП» и, следовательно, уменьшать ошибку измерения, обусловленную механическими свойствами кнопки.
В качестве монохроматического источника света использовался RGB-светодиод (Ultra brightness BL-L515RGBC) с рассеивающей линзой (угол рассеивания 200), что позволяло инициировать вспышку красного (Ultra Orange 630 nm), зеленого (Ultra Pure Green 525 nm) или синего (Blue 430 nm) света. Значение силы света для каждого излучателя (красного, зеленого и синего) составляло постоянную величину и аппаратными средствами устанавливалось на приемлемом для глаза уровне 2000 mcd (luminous intensity). Световой излучатель диаметром 5 мм размещался в центре квадратной подложки белого цвета (80 х 80 мм).
Статистический анализ проводился с использованием пакетов прикладных программ Excel и Statistica 7.0 путем расчета основных показателей описательной статистики. Статистическую значимость различий между двумя средними арифметическими величинами определяли с помощью двухвыборочного t-теста Стьюдента для выборок с различными дисперсиями при заданном уровне значимости p<0,05. Для оценки характера зависимости между исследуемыми параметрами использовались регрессионный и корреляционный анализ. Для характеристики вариабельности исследуемых показателей рассчитывались величины дисперсии выборок и коэффициенты вариации. Во всех случаях сравнение анализируемых показателей и оценка статистической значимости различий проводились на основании проверки нулевой и альтернативной гипотез.
Результаты
В эксперименте № 1определялись условия формирования полной обратной световой маскировки: длительность тестового стимула и стимула-маскера, а также «критическое время асинхронии» (АВСкрит). Результаты исследования показали, что эффект полной обратной световой маскировки проявляется при длительности ТС = 0,1 мс, МС = 30 мс и значении АВС-интервала в диапазоне от 0 до 15 мс (табл. 1). При таких условиях опыта все испытуемые в 100% случаев уверенно отвечали: «Вижу только зеленый свет», т.е. стимул-маскер (Зеленый) полностью маскировал тестовый стимул (Синий). При дальнейшем удлинении АВС-интервала до 20, 25 и 30 мс эффект полной маскировки наблюдался соответственно в 79 ± 6,1%, 63 ± 4,6% и 13 ± 4,0% случаев (p< 0,05). Наконец, при АВС-интервале, равном 40 мс, все испытуемые и во всех пробах отвечали: «Вижу синий и зеленый стимулы». Такое состояние свидетельствовало об исчезновении эффекта полной маскировки, а, следовательно, интервал АВС = 40 мс соответствовал «критическому времени асинхронии включения стимула» АВСкрит..
Таблица 1
Динамика развития эффекта полной обратной световой маскировки двухцветной модели (M ± m) при разных значениях параметров стимуляции
|
Условия эксперимента № 1 |
|
|
Параметры стимуляции (ТС — МС — АВС), мс |
Показатель степени полной обратной маскировки, % |
|
0,1 — нет — нет |
- |
|
0,1 — 0,1 — 0 |
0 |
|
0,1 — 30 — 0 |
100 |
|
0,1 — 30 — 10 |
100 |
|
0,1 — 30 — 15 |
100 |
|
0,1 — 30 — 20 |
79 ± 6,1 |
|
0,1 — 30 — 25 |
63 ± 4,6 |
|
0,1 — 30 — 30 |
13 ± 4,0 |
|
0,1 — 30 — 40 |
0 |
Примечание. ТС — тестовый стимул (Синий); МС — маскер-стимул (Зеленый); АВС—асинхрония включения стимулов.
В эксперименте № 2 определялось время формирования физиологического феномена как составной части латентного периода дифференцировочной зрительно-моторной реакции (табл. 2, рис. 1). С этой целью время ДЗМР (RT + МС) измерялось на фоне создания помехи исследуемой сенсомоторной реакции с помощью стимула-маскера (МС), предъявляемого испытуемому при разном значении АВС-интервала. Следует напомнить, что в данной серии опытов стимул-маскер предъявлялся в период, следующий после исчезновения эффекта полной обратной маскировки, т. е. начиная с 40 мс. Как видно из табл. 2, при значении АВС-интервала 40 мс, соответствующего для всей группы испытуемых «критическому времени асинхронии включения стимулов, АВСкрит», латентный период зрительно-моторной реакции был на 106 ± 11,9 мс более продолжительным, чем в условиях без предъявления сенсорной помехи. В дальнейшем, вместе с удлинением АВС-интервала, латентный период при выполнении ДЗМР неуклонно укорачивался и при АВС = 300 мс уменьшался до 340 ± 6,3 мс, т. е. статистически значимо не отличался от величины, регистрируемой в условиях «без создания помехи» (343 ± 5,6 мс). Коэффициенты вариации во всех случаях являлись невысокими и сравнимыми по величине.
Алгоритм проведения теста позволил определить величину АВС-интервала, начиная с которой эффект помехи на латентный период исследуемой сенсомоторной реакции более не проявлялся (табл. 2, рис. 1). Так, при значении показателя «время асинхронии включения стимулов», равного 100 мс, латентность дифференцировочной зрительно-моторной реакции составила 347 ± 6,6 мс, т. е. соответствовала уровню в опыте «без создания помехи» (RT, 343 ± 5,6 мс). Оценка степени различий между (RT+МС) и RT подтвердила сделанный вывод о том, что при удлинении АВС-интервала до 100 мс влияние стимула-маскера на характер и скорость зрительно-моторной реакции не обнаруживается; в этом случае различия составили 4,0 ± 8,6 мс и являлись статистически незначимыми (p>0,05). Следовательно, можно предположить, что к этому моменту времени, т. е. через 100 мс от начала момента предъявления тестового стимула, фаза формирования физиологического феномена, как составной части латентного периода дифференцировочной зрительно-моторной реакции, завершалась.
Таблица 2
Значение латентного периода дифференцировочной зрительно-моторной реакции (RT) при разных условиях эксперимента
|
Значение АВС, мс |
Условия эксперимента и показатель |
||
|
Латентный период дифференцировочной зрительно-моторной реакции в опытах с созданием помехи стимулом-маскером (RT + MС), мс |
Латентный период дифференцировочной зрительно-моторной реакции в опытах без создания помехи стимулом-маскером (RT), мс |
Разница между (RT + MС) и RT, мс |
|
|
40 |
449 ± 10,6*(1346)[8,2%] |
343 ± 5,6 (376) [5,7 %] |
106 ± 11,9 ** |
|
50 |
431 ± 8,3*(823)[6,7%] |
88 ± 10,0 ** |
|
|
60 |
405 ± 7,7*(708)[6,6%] |
62 ± 9,5 ** |
|
|
70 |
383 ± 7,2*(620)[6,5%] |
40 ± 9,1 ** |
|
|
80 |
364 ± 6,7*(538)[6,4%] |
21 ± 8,7 ** |
|
|
90 |
361 ± 6,3*(475)[6,0%] |
18 ± 8,4 ** |
|
|
100 |
347 ± 6,6(520)[6,6%] |
4 ± 8,6 |
|
|
150 |
346 ± 7,1(600)[7,1%] |
3 ± 9,0 |
|
|
200 |
349 ± 6,1(445)[6,0%] |
6 ± 8,2 |
|
|
300 |
340 ± 6,3(475)[6,4%] |
-3 ± 8,4 |
|
Примечание. «*»—различия по отношению к показателю RT «без создания помехи» статистически значимы (p<0,05); «**»—значение вычисленной разницы в показателях статистически значимо (p<0,05); в круглых скобках приведено значение дисперсии выборки; в квадратных скобках приведены коэффициенты вариации.
Собственно весь процесс (рис. 1, А) во всем исследованном диапазоне АВС-интервалов (40—300 мс) лучше всего описывался гиперболической зависимостью следующего вида: y = 310 + (5438 / x) при статистически значимом коэффициенте детерминации (R2 = 0,92) и небольшой средней ошибке аппроксимации 2,42%.
Для более полной характеристики зависимости между латентным периодом ДЗМР и значением АВС-интервала был проведен соответствующий корреляционный анализ, во-первых, в более узком диапазоне значений асинхронии включения стимула и, во-вторых, с использованием логарифмической шкалы (рис. 1, Б). При этом учитывался тот факт, что эффект задержки скорости выполнения ДЗМР при предъявлении стимула-помехи наблюдался в диапазоне АВС от 40 до 90 мс, а при АВС, равном 100 мс, эффект полностью исчезал. Результаты анализа показали, что в указанном диапазоне значений АВС (40—100 мс) в логарифмической шкале характер зависимости описывается уравнением прямой линии следующего вида: y = -116 (х) + 877 при высоком коэффициенте детерминации 0,99 и небольшой его ошибке (1,05%). Значение всех коэффициентов в уравнении регрессии были статистически значимы (p<0,05). Такой результат свидетельствует о том, что латентный период (физиологический эффект, «у») пропорционален логарифму АВС-интервала («х»), что, по сути, соответствует критериям основного психофизического закона Вебера—Фехнера, согласно которому сила ощущения пропорциональна логарифму интенсивности раздражителя. Следует принять во внимание, что в нашем случае было показано: чем меньше латентный период, тем больше скорость сенсомоторной реакции, или тем более выражен физиологический эффект.
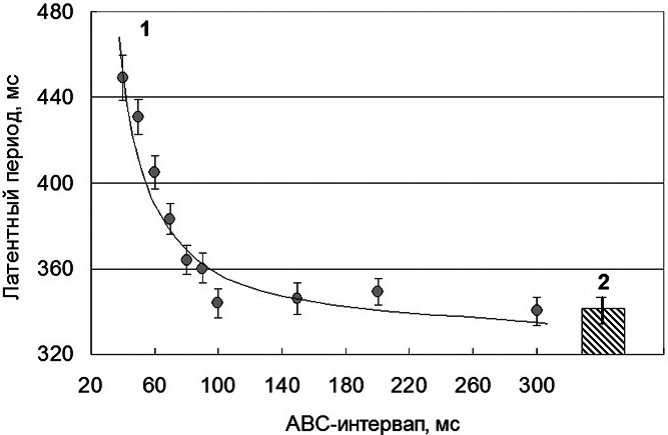

Рис. 1. Характер зависимости латентного периода дифференцировочной зрительно-моторной реакции «Go/No-go» – типа от значения АВС-интервала в условиях создания сенсорной помехи путем дополнительного предъявления стимула-маскера в арифметической (А) и логарифмической (Б) шкалах: на А—значение АВС-интервала приведено в арифметической шкале во всем исследованном диапазоне АВС (от 40 до 300 мс); 1 — теоретическая кривая гиперболической зависимости, 2 — значение латентного периода в условиях без предъявления стимула-маскера, мс; на Б—значение АВС-интервала (от 40 до 100 мс) приведено в логарифмической шкале
Эксперимент № 3. На заключительном этапе исследования осуществлялся анализ экспериментальных данных по результатам тестирования испытуемых на предмет субъективной оценки степени выраженности ощущения при предъявлении тестового стимула на фоне действия стимула-маскера (помехи) при разных значениях АВС-интервала. Анализ показал, что при значении АВС-интервала 40 мс средняя оценка степени выраженности светового ощущения составила 1 балл (при RT = 449 ± 10,6), а при 70 и 100 мс — 2,17 ± 0,11 (при RT = 383 ± 7,2) и 2,92 ± 0,08 (при RT = 347 ± 6,6) балла соответственно. Эти результаты свидетельствуют о том, что: а) запуск выполнения дифференцировочной зрительно-моторной реакции осуществляется во всех случаях, несмотря на существенно различающуюся степень выраженности светового ощущения; б) между интенсивностью ощущения и латентным периодом ДЗМР существует отрицательная корреляционная связь с коэффициентом корреляции Пирсона -0,99 ± 0,05 (p< 0,01).
Обсуждение результатов
Реализация дифференцировочной зрительно-моторной реакции (ДЗМР), с позиций системного подхода П.К. Анохина [Анохин, 1978], осуществляется с помощью специфической функциональной системы, которая была сформирована и помещена в память ранее на основании индивидуального опыта, но в искомый момент времени после постановки перед испытуемым экспериментальной задачи извлечена из памяти, активирована и скорректирована после нескольких пробных запусков. При этом в качестве аксиомы принимается принципиально важное допущение: реализация зрительно-моторной реакции «Go/No-go» типа предполагает непременное опережающее моторный эффект формирование у испытуемого осознанного ощущения цвета тестового и тормозного стимулов.
Для реализации ДЗМР необходимы, по крайней мере, два важных условия — формирование физиологической и психической компоненты ощущения. Изначально формируется физиологический феномен (ФФ), который при ДЗМР «Go/No-go» типа сам по себе не запускает моторную реакцию, так как характер цвета тестового и тормозного стимулов к этому моменту еще не осознан. Непосредственный запуск ДЗМР возможен только после распознания цвета стимула, т.е. формирования психического феномена ощущения. В парадигме обратной световой маскировки фаза формирования ФФ характеризуется двумя знаковыми точками, располагающимися в рамках диапазона АВС-интервала: АВСкрит и АВСфф.. При наших условиях экспериментов среднее значение АВСкрит в группе испытуемых составило 40 мс. К этому времени сенсорная помеха в виде стимула-маскера у всех испытуемых полностью утратила способность блокировать процесс последующего осознания предъявленного тестового стимула. Величина АВСкрит может колебаться в широком диапазоне значений и определяется условиями обратной маскировки [Бахманн, 1989; Соболев В.И, 2020].
Измерение длительности ФФ в наших экспериментах проводилось путем нахождения интервала времени (АВСфф) между тестовым стимулом и стимулом сенсорной помехи (маскера), при котором сенсорная помеха, предъявляемая на разных этапах реализации зрительно-моторной реакции «Go/No-go» типа, полностью утратит способность замедлять скорость последней. Предполагалось, что к окончанию этого периода завершится формирование физиологического феномена, который в соответствующей форме транслируется в следующие структурные элементы центрального звена реализации зрительно-моторной реакции, и попытки помешать этому процессу становятся невозможными. По нашим данным, такой интервал, обозначенный как АВСфф, составляет не менее 100 мс, так как после этой временной точки сенсорная помеха (маскер) была уже не в состоянии что-либо изменить в реализации психомоторной реакции. Как мы полагаем, интервал АВСфф длительностью 100 мс необходим и достаточен при данных условиях эксперимента для полноценного формирования физиологического феномена как психофизической основы последующего проявления осознанного ощущения (психического феномена).
Найденное значение АВСфф (100 мс) облегчает поиск места локализации нейрональной структуры, в которой возможно завершение процесса формирования ФФ или, по крайне мере, его наиболее важных, пока еще неизвестных, составляющих. По-видимому, эти структуры должны располагаться на временной шкале в точке на расстоянии не более 100 мс от момента предъявления тестового стимула. С учетом сказанного можно предположить, что точка завершения процесса формирования ФФ топографически может быть локализована в одном из пунктов «кольца» А.М. Иваницкого [Иваницкий, 1996] либо в функционально подобных ему образованиях, описанных В.Я. Сергином [Сергин, 2016], G. Edelman [Edelman, 2000] и др. Действительно, в предложенной А.М. Иваницким схеме «кольца» весь цикл кольцевого движения возбуждения занимает 150 мс [Иваницкий, 1996] без учета еще 30 мс, необходимых для поступления возбуждения от сетчатки в первичную зрительную кору (вход «кольца»). Если за точку отсчета принять момент предъявления испытуемому светового стимула, то через 100 мс возбуждение достигнет зрительных ассоциативных зон (поля Бродмана 18 и 19), т. е. структуры, где, как мы предполагаем, завершается процесс формирования полноценного ФФ. Это значение практически совпадает с эмпирически найденным нами интервалом АВСФФ 100 мс. Полагают, что именно в экстрастриарной коре преимущественно осуществляется цветовое декодирование стимула с участием структур с цветооппонентными характеристиками [Измайлов, 1989; Шиффман, 2003]. В дальнейшем, сформировавшийся физиологический феномен или его отдельные ключевые компоненты в виде нейрофизиологического паттерна, содержащего всю информацию о цветовом стимуле, достигает гиппокампа или в целом гиппокамповой формации, а далее движется по «кольцу Иваницкого». Детали этого процесса нуждаются в конкретизации, которая может быть осуществлена с помощью прямых методов нейрофизиологического исследования, в связи с чем, предлагаемая нами схема, равно как и собственно трактовка понятия физиологического феномена, носят дискуссионный характер.
Второй аспект исследования, представляющий интерес для обсуждения, связан с тем, что предъявление стимула-помехи (маскера) на участке диапазона «АВСкрит.—АВСФФ», т. е. от 40 до 100 мс, не блокировало последующее формирование осознанного ощущения и, соответственно, реализацию ДЗМР, однако оказывало выраженное отрицательное влияние на латентность зрительно-моторной реакции. Так, латентный период ДЗМР при данных условиях максимально удлинялся на 106 ± 10,6 мс (табл. 2, АВС = 40 мс), но при АВС = 100 мс эффект торможения реакции у всех испытуемых полностью исчезал. Это можно объяснить завершением к этому моменту времени периода формирования ФФ и «ускользанием» процесса реализации ДЗМР от тормозящего действия стимула-помехи.
В контексте обсуждения механизма действия стимула-маскера принципиально важным обстоятельством представляется обнаруженная зависимость между латентностью дифференцировочной зрительно-моторной реакции (у) и значением АВС-интервала (х), которая описывается в логарифмической шкале прямой линией (рис. 1, Б). Если принять, как аксиому, что скорость ДЗМР в конечном итоге эквивалентна интенсивности ощущения, а АВС-интервал отражает интенсивность раздражителя, то указанная зависимость в точности соответствует критериям известного закона Вебера—Фехнера, согласно которому сила ощущения пропорциональна логарифму интенсивности раздражителя. Различия заключаются лишь в том, что в законе Вебера—Фехнера интенсивность раздражителя характеризуется физическими параметрами тестового стимула (яркость, длительность светового стимула и др.), тогда как в нашем случае — интенсивностью нейронального ответа в виде физиологического феномена, сформировавшегося в ходе анализа информации о стимуле и эквивалентного длительности АВС-интервала. Таким образом, можно полагать, что замедление скорости ДЗМР происходит не за счет временной задержки процесса в нейрональных структурах «кольца» А.М. Иваницкого или подобных ему образованиях, а за счет уменьшения (или искажения) под влиянием стимула-маскера (помехи) силы (интенсивности, структуры) формирующегося физиологического феномена либо его отдельных ключевых компонентов. В результате на выходе «кольца» появляется спектр ослабленных сигналов о тестовом стимуле различной интенсивности, формирующих в дальнейшем на уровне восприятия психические феномены разной степени выраженности и, как следствие, инициирующих большую или меньшую скорость сенсомоторной реакции, что мы и наблюдали.
Такой вывод согласуется и с результатами 3-й серии экспериментов с использованием метода анкетирования испытуемых: субъективная оценка степени выраженности осознанного ощущения тестового стимула отрицательно коррелировала с длительностью АВС-интервала. В связи с этим можно выстроить цепочку, отражающую особенности механизма действия маскера-помехи на латентный период ДЗМР: укороченный АВС-интервал (при условии: АВСкрит<АВС<АВСФФ) — ослабленный физиологический феномен — слабо выраженное осознанное ощущение — замедление скорости ДЗМР.
Все сказанное может рассматриваться как подтверждение гипотезы о том, что механизм торможения ДЗМР при обратной маскировке вследствие действия сенсорной помехи, предъявляемой на стадии формирования ФФ, связан первоначально с модуляцией интенсивности либо с нарушением конфигурации физиологического феномена в структурах, подобных «кольцу» А.М. Иваницкого, а затем и выраженностью формируемого осознанного ощущения (психического феномена).
Выводы
- Эффект полной (100%) обратной световой маскировки (ОМС) надежно реализуется при следующих значениях ее параметров: длительность тестового стимула (Синий) 0,1 мс, длительность стимула-маскера (Зеленый) 30 мс, диапазон асинхронии включения стимулов (АВС) 0—15 мс; при АВС-интервалах 20, 25 и 30 мс степень проявления полной маскировки в группе испытуемых последовательно снижается до 79±6,1, 63±4,6 и 13±4,0%, а при АВС, равной 40 мс, полностью исчезает (0%).
- Выявлен эффект торможения дифференцировочной зрительно-моторной реакции (ДЗМР) «Go/No-go» – типа под влиянием стимула-маскера, предъявляемого в разные периоды ее реализации (ReactionTime, RT); такой эффект проявляется в диапазоне АВС от 40 до 100 мс; между RT и значением АВС-интервала в указанном диапазоне существует зависимость, описываемая в логарифмической шкале уравнением прямой линии: латентность сенсомоторной реакции обратно пропорциональна логарифму АВС.
- Для измерения длительности периода формирования физиологического феномена, как предиктора осознанного ощущения, может быть использован психофизиологический метод, основанный на измерении степени торможения скорости дифференцировочной зрительно-моторной реакции «Go/No-go» типа, реализуемой в парадигме обратной световой маскировки; установлено, что латентность формирования физиологического феномена составляет 100 мс.
- Возможный механизм торможения скорости выполнения ДЗМР «Go/No-go» типа при обратной маскировке заключается в ослаблении под влиянием стимула-маскера интенсивности формирующегося физиологического феномена, что в конечном итоге закономерно вызывает удлинение латентного периода сенсомоторной реакции.