Введение
В общем объеме времени, которое человек затрачивает на поиск, обнаружение и опознание значимых элементов среды, фиксации занимают 80—85%. Роль саккад (15—20% времени восприятия) как компонента перцептивного процесса, несмотря на то, что их продолжительность также сопоставима с проявлениями значимых событий в среде, остается неопределенной. Чаще всего это связывают с наличием «саккадического подавления» — резким снижением зрительной способности, сопровождающим быстрые движения глаз, а на уровне теории — с представлением о дискретности приема и переработки зрительной информации, активная фаза которых реализуется во время фиксации.
Эксперименты показывают, что саккадическое подавление действительно существует, но не является биологической константой и зависит от различных обстоятельств. В ходе резких поворотов глаз пороги зрительной чувствительности повышаются частично и зависят от амплитуды саккады (с ее ростом чувствительность падает), пространственной структуры тестового стимула (при наличии выраженных контуров подавление усиливается), его яркости (с увеличением освещенности тест-объекта эффект подавления также усиливается) и локализации в поле зрения (пороги зрительного обнаружения стимула распределены анизотропно: зона максимальной чувствительности соотносится с текущим объектом фиксации, а при повороте глаз смещается в область будущей фиксации). Хотя усложнение тестового стимула (его структурированность, контрастность и др.) приводит к усилению эффекта подавления, возможность получения семантической информации об объекте во время саккад сохраняется [Brooks, 1975; Campbell, 1978; Cavanagh, 2010; Honda, 1989; Mitrani, 1975; Ross, 2001; Schlag, 1995]. С увеличением размеров стимула, а также перепада яркости «фигура—фон» подавление растет. Структурированный фон, детализированная текстура повышают саккадические пороги чувствительности [Brooks, 1975]. Mitrani, Yakimoff, Mateeff (1973) нашли, что подавление возникает, когда структурированный фон присутствует в начале саккады. Наличие его на остальных участках движения не оказывает влияния. Это связывают с тем, что структурированный, стабильно воспринимаемый фон перед саккадой выступает контрастом для тест-объекта, предъявленного строго интра-саккадически, и инициирует эффект прямой зрительной маскировки. Заход экспозиции объекта за временные границы саккады нарушает этот эффект. Чем более структурирован объект, тем меньше он маскируется неструктурированной маской [Ellis, 1971; West, 1968]. Низкие частоты подавляются максимально сильно [Burr, 1994]. Из временами противоречивых выводов, сделанных разными авторами в разное время, можно предположить, что эффективность восприятия, сопровождающего движения глаз, зависит не от пространственной частоты тест-объекта или фона самих по себе, а от их соотношения. Более того, при одних условиях на подавление влияет яркость, а при других — наличие четких контуров (структуры) [Mitrani, 1975].
Отсутствие метаконтраста при условии постсаккадического исчезновения (temporal gap) тест-объекта зафиксировали Deubel, Schneider, Bridgeman (1996). Данный факт не противоречит принципу необратимости зрительного процесса, поскольку речь идет о проявлении ранее воспринятой информации, а не о ее восстановлении. Наличие близлежащих дистракторов в фовеальной зоне усиливает восприятие контраста, в периферической — ослабляет, причем маскер, совпадающий с основным объектом по пространственной частоте и ориентации линий, оказывает более сильный эффект [Petrov, 2005]. Большая часть выполненных работ регистрирует саккадическое подавление в диапазоне от -80 мс (до начала саккады) до +50 мс (после начала саккады).
Классические представления о саккадическом подавлении сформировались в 60—70-х гг. прошлого века по результатам экспериментальных исследований зрительной чувствительности к геометрическим примитивам во время саккадических движений глаз [Kowler, 2011; Latour, 1962; Matin, 1974; Mitrani, 1973; Volkmann, 1986]. В качестве стимулов использовались точки, вспышки света, пространственные решетки. В последние два десятилетия было отмечено несколько работ с применением натуральных стимулов [Dorr, 2013], обладающих экологической валидностью. Однако ни в одной из них не исследовались феномены интра-и парасаккадического подавления в отношении лицевых экспрессий.
Наши работы по динамике зрительной способности во время саккад проводятся в русле проблем восприятия экологически или социально значимых событий. В центре внимания — распознавание и использование лицевых экспрессий коммуникантов в ходе межличностных взаимодействий. Наряду с психофизическими характеристиками зрительного процесса особая роль отводится ценностному отношению к предмету восприятия, его значению в контекстах общения и совместной деятельности [Барабанщиков, 2016; Барабанщиков, 2014; Barabanschikov, 2019; Zherdev, 2020]. В серии экспериментов изучалось распознавание базовых экспрессий лица во время выполнения целенаправленной саккады; в качестве дополнительной процедуры определялось воспринимаемое местоположение лица в поле зрения. Оценка модальности экспрессий осуществлялась путем выбора из двух альтернатив. Оказалось, что средняя частота верного распознавания экспрессий при средней скорости саккады 225°/с выше случайной и равна 0,61. Это значение кратно превышает частоту обнаружения в сходных условиях точечных вспышек света или распознавания геометрических фигур [Latour, 1962; Mitrani, 1973; Volkmann, 1962], но на 15—20% меньше точности опознания эмоциональных выражений лица в условиях свободного рассматривания. Лучше всего в данной ситуации распознаются проявления радости (0,81) и страха (0,71), сравнительно плохо — «гнев» (0,54) и «печаль» (0,56); хуже всего — «спокойное состояние» (0,43). В силу сходства паттернов экспрессий устойчиво путаются «радость» и «страх», «гнев» и «отвращение», «печаль» и «гнев» и др.
Выбор подходящей эмоции, который делает наблюдатель, обусловлен как модальностью экспрессии, экспонированной во время саккады, так и модальностью альтернативной экспрессии, с которой сравнивается тест-объект. Результат выбора зависит от аттрактивности (визуальной броскости) выражений лица. Если экспрессия выступает в роли тест-объекта, высокая аттрактивность содействует адекватному выбору, если же она экспонируется в качестве альтернативы — выбор затрудняется. Чем лучше распознается базовая эмоция, тем больший маскирующий эффект она может вызвать.
Перцептивный переходный процесс, объединяющий смежные фиксации, сопровождается ложной локализацией стимулов [Honda, 1991; Matin, 1965] и их сжатием в направлении выполняемой саккады [Burr, 2010; Lappe, 2000]. Независимо от того, где во время саккады находится изображение лица, наблюдатели локализуют его в одних и тех же узких участках поля зрения (0,6° в правой части, 1,8° — в левой), прилегающих к будущей точке фиксации. Нарушений константности зрительного направления (стабильности видимого мира) в подавляющем большинстве случаев не происходит.
При переводе взора с одного точечного объекта на другой выделяются три фазы зрительного процесса, последовательно сменяющие друг друга: 1) при фиксации креста в центре экрана непосредственно (-80 — 0 мс) до выполнения саккады средняя точность распознавания экспрессии соответствует точности распознавания выражения лица, расположенного на периферии на расстоянии 10°; 2) во время саккады (средняя длительность 48 мс) точность опознания плавно растет; 3) в начале новой фиксации (48—180 мс от начала саккады) высокая эффективность распознавания достигает максимума и стабилизируется. Разрывов либо резких падений зрительной способности в парасаккадическом интервале не выявлено. Перед началом саккады отмечены слабая тенденция к снижению уровня идентификации (интервал от -30 до -15 мс) и повышенная вариативность ответов.
Экспериментальные данные свидетельствуют о том, что тактовая структура окуломоторной активности (фиксация—саккада—фиксация) не нарушает пространственно-временную динамику зрительного процесса. Перцептогенез выражения лица совершается не только в период устойчивой фиксации взора, но и на пике скорости быстрых движений глаз (около 400°/с), причем как в центре, так и на ближней периферии (±10° от центральной точки фиксации) зрительного поля. В процессе распознавания сложных социально значимых объектов имеет место не столько подавление зрительной способности, как некоего самостоятельного механизма организации перцептивного процесса, сколько усредненное снижение эффективности предметного восприятия, вызванное эксцентрическим расположением лица.
Результаты проведенных экспериментов говорят о том, что мы столкнулись с группой перцептивных явлений, несколько отличной от той, которую принято обозначать термином «саккадическое подавление». Несмотря на процедурное сходство — экспозицию кратковременных стимулов в ходе выполнения саккад, в парадигме наших исследований (1) используются не абстрактные, а натуральные стимульные объекты, имеющие для человека жизненно важное значение; (2) вводятся характеристики активности воспринимающего; (3) наблюдатели выполняют задачу опознания (идентификации), а не поиска и обнаружения; (4) процедура не ограничивается искусственно выделенным фрагментом зрительного процесса, связанным с исполнением быстрых движений глаз, и учитывает целостность перцептивного события (его эпизода). Основным предметом исследования становится процесс взаимодействия субъекта с объектом, протекающий в микроинтервалах времени, который опирается на информацию, заключенную в предшествующей фиксации, и реализуется в последующей. Мы называем его транссаккадическим восприятием. В ходе исследования делается акцент не на подавление зрительной способности, а на ее динамике в различные моменты времени.
В новой работе экспериментально изучалось влияние предсаккадического стимула на восприятие интрасаккадического и сопоставлялась динамика идентификации экологически валидных объектов с оценками элементарных оптических свойств и отношений, выполненных в сходных режимах экспозиции; собирались дополнительные аргументы, подтверждающие результаты более ранних исследований транссаккадического восприятия лица.
Метод исследования
Идея эксперимента состоит в том, чтобы разделить и противопоставить экологически валидные элементы одной и той же стимульной ситуации при переводе взора наблюдателя на новый объект. Имеется в виду создание условий параконтраста, когда содержание одного (латерального) стимула, становясь предметом внимания, инициирует саккаду, а тестовый стимул, несущий другое, но однородное содержание, появляется во время целенаправленных движений на месте первого. Пара-и интрасаккадическая информация оказываются здесь в одном и том же зрительном поле, позволяя исследовать характер их отношений, в том числе обстоятельства, меняющие зрительную способность на различных этапах перцептивного переходного процесса [Барабанщиков, 2002].
Оборудование. Экспериментальная установка состояла из айтрекера SMI HiSpeed-1250 и стимульного компьютера со специализированным программным обеспечением.
Процедура. Перед испытуемыми ставилась задача в каждой пробе перевести взгляд с фиксационного креста в центре экрана на латеральный объект сразу после его появления. С помощью прямоугольной рамки, высвечиваемой на экране, требовалось обозначить видимую локализацию стимула — изображение лица, переживающего эмоциональное состояние, а затем идентифицировать модальность эмоции. Озвученная инструкция побуждала к выполнению реактивной саккады и не позволяла заранее понять, когда именно должен экспонироваться тест-объект и от чего это зависит.
В начале пробы появлялся фиксационный крест. Программное обеспечение проверяло устойчивость фиксации взора на кресте; через 3000 мс, фиксационный крест сменялся латеральным объектом, смещенным относительно центра экрана на 10° влево или вправо (рис. 1).
Роль латерального объекта выполняло одно из изображений экспрессии (отличной от тест-объекта, экспонируемого интрасаккадически). Начало саккады инициировало одновременно появление тест-объекта (всегда на 1 кадр развертки монитора, длительностью 6,92 мс) и исчезновение латерального стимула (рис. 2). Точный момент начала и завершения всех этапов каждой пробы регистрировался посредством фотодиодного сенсора, прикрепленного на экране. Через 100 мс после исчезновения тест-объекта появлялась рамка, необходимая для выполнения задачи на локализацию, а затем — варианты ответа, необходимые для идентификации экспрессии (случайные позиции друг относительно друга).
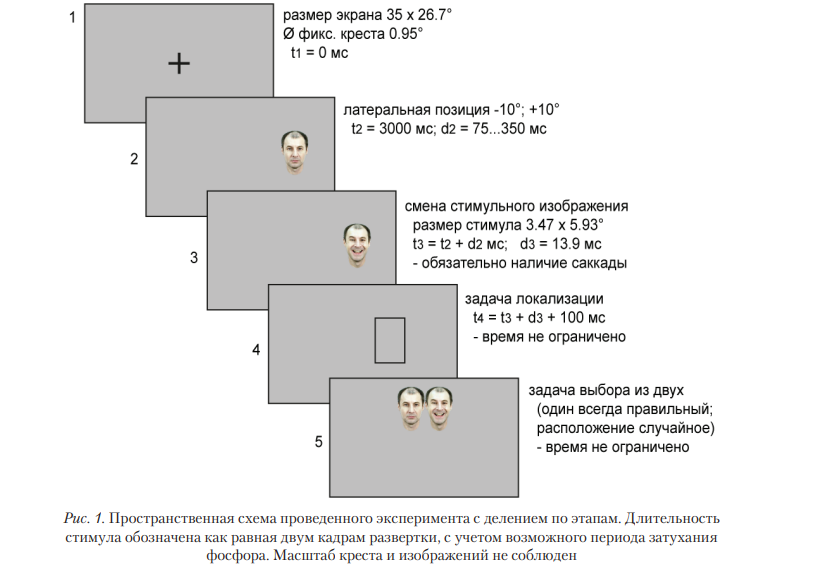
Временная развертка событий, протекающих в каждой пробе, представлена на рис. 2. Описанная процедура напоминает парадигму temporal gap, но роль стимула выполняет изображение, а не исчезновение объекта, и стимул сменяется во время саккады. Оценка локализации и распознавания выполнялись при помощи движения и нажатия кнопки компьютерной мыши. Использование клавиатуры не требовалось. Продолжительность выполнения заданий не ограничивалась.
Стимульный материал. В случайном порядке экспонировались цветные фотоизображения мужского лица, выражающего шесть базовых экспрессий + спокойное состояние из фотобазы ВЕПЭЛ [Куракова, 2012]. Максимальная яркость — 79,8 кд/м2. Угловой размер изображений — 3,47 x 5,93° зрительного угла. Логарифм относительной яркости фигуры-фона = 0,24. Цвет фона — 75% серый, яркостью 46,3 кд/м2. Доминантная пространственная частота 2 cpd (цикла на градус).
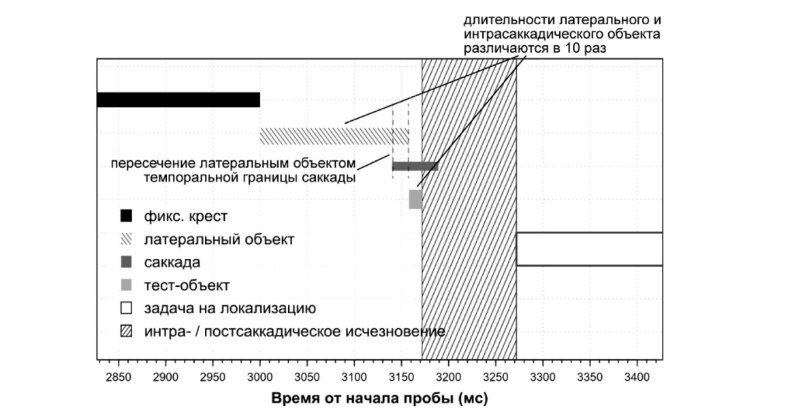
Анализ пространственной частоты фотоизображений выполнен посредством нахождения габоровских решеток для фильтра различного размера и ориентации. На рис. 3 приведены исходные изображения, под каждым из них — габоровские решетки в четырех ориентациях. Поскольку от испытуемого требовалось выполнение горизонтальной саккады, то пространственная частота посчитана по решетке с вертикальной ориентацией фильтра (см. фрагмент 2). Из рисунка видно, что горизонтальная пространственная частота для всех экспрессий равна 7 cpf (цикла на уровне глаз), что с учетом углового размера стимула в эксперименте соответствует 2 cpd (циклам на градус).
Обработка данных. Перед началом статистической обработки данные были подвергнуты фильтрации на основании заданных критериев экспериментального протокола: латентность реактивной саккады должна быть в диапазоне 75—350 мс от появления латерального объекта; позиция взора в момент начала экспозиции тест-объекта должна находиться в пределах от 1,85° до 8,15° (на пути следования саккады с амплитудой 10° зрительного угла); длительность тест-объекта должна быть равна 1—2 кадрам вертикальной развертки монитора. Статистической обработке подверглись 233 (12%) валидные пробы.
Использовались непараметрический критерий Хи-квадрат, тест Колмогорова— Смирнова — для проверки нормальности, для сравнения метрических данных — t-тест. Все тесты — двусторонние. Статистическая обработка выполнена в среде R, версия 4.0.2.
Выборка испытуемых. В исследовании приняли участие 21 человек (9 женщин) с нормальным зрением. Медианный возраст — 22 ± 6.
Результаты
Окуломоторные показатели. При выполнении перцептивной задачи зарегистрированы следующие показатели целенаправленных саккад: латентность саккады — 140±57 мс; длительность саккады — 46±11 мс; амплитуда — 8,6±1,3°; средняя скорость саккады — 183±36 °/с; пиковая скорость — 326±50 °/с. Существенно, что латентность саккады к латерально расположенной лицевой экспрессии меньше, чем к латерально расположенному миниатюрному кресту в сходной стимульной ситуации [Barabanschikov, 2019]; t-тест = 9,56(430) (здесь и далее в скобках указано количество степеней свободы); p<0,001; d Коэна = 0,91 (большой размер эффекта); значение мощности — 0,53.
Идентификация. Корректной идентификации интрасаккадического тест-объекта не наблюдается (точность идентификации — 22%), тест Хи-квадрат = 72,5(1); p < 0,001. Вместе с тем испытуемые надежно идентифицируют латеральный объект (точность — 78%). Метрика F1 для частоты верной идентификации приведена на рис. 4. Экспрессия тест-объекта связана с корректной идентификацией, тест Хи-квадрат = 32,96(6); p < 0,001; экспрессия альтернативы (N.B. Мы условно полагаем, что латеральный объект — альтернатива, интрасаккадическая экспозиция — правильный вариант ответа) также связана с идентификацией, тест Хи-квадрат = 47,74(6); p < 0,001. Стоит отметить, что испытуемые получали инструкцию с задачей ‘обозначить ту экспрессию, которую вы видели’. В ней не было указано, что она будет сменяться во время саккады, или каких-либо упоминаний о первой и второй экспрессии. Испытуемые были поставлены в ситуацию свободной интерпретации предъявляемых стимулов.
Если проводить аналогию с работой Deubel, Schneider, Bridgeman (1996) и принимать во внимание медианную латентность саккады в данном эксперименте, равную 140 мс, то результаты нашего исследования согласуются по частоте идентификации первого стимула (латерально смещенной цели). Авторы нашли, что минимальная частота идентификации второго тест-объекта (интрасаккадического) должна быть при околонулевом временном промежутке между первым и вторым стимулами. Это условие выполняется в нашей процедуре, за исключением того, что латеральный объект имеет намного более длительную экспозицию. При латентности в 140 мс, согласно указанным авторам, частота идентификации равна 82%. В нашем эксперименте это условие не выполняется только для экспрессии радости.
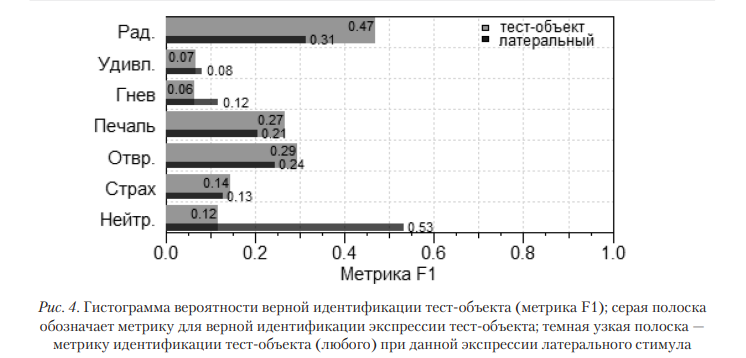
Относительность оценок. Из данных, приведенных на рис. 4, следует, что для экспрессий с самыми низкими уровнями идентификации (широкие полосы на графике) интеграционный процесс практически отсутствует, а интрасаккадический тест-объект подвержен маскировке. Даже вероятность идентификации экспрессии радости остается на уровне шанса. Чем лучше распознается базовая эмоция, тем эффективнее ее маскировка. Нейтральная экспрессия в роли альтернативного варианта (в качестве латерального объекта) усиливает корректную идентификацию; остальные экспрессии обладают смешанным эффектом. Так, экспрессии удивления и гнева предпочитаются испытуемыми в качестве ответа, когда они экспонированы в качестве латерального объекта (обладают высокой визуальной аттрактивностью). Однако они не идентифицируются в роли стимула. Для успешной идентификации требуется несколько факторов, среди которых модальность экспрессии и длительность ее экспозиции (для разных экспрессий требуется различная длительность). Более того, их базовые уровни могут быть значимо ниже, чем теоретическое 0,5. Это говорит о наличии некоторого фактора (группы условий), который не просто затрудняет идентификацию, но и снижает ее до необычно низкого уровня, значимо ниже вероятности случайного угадывания.
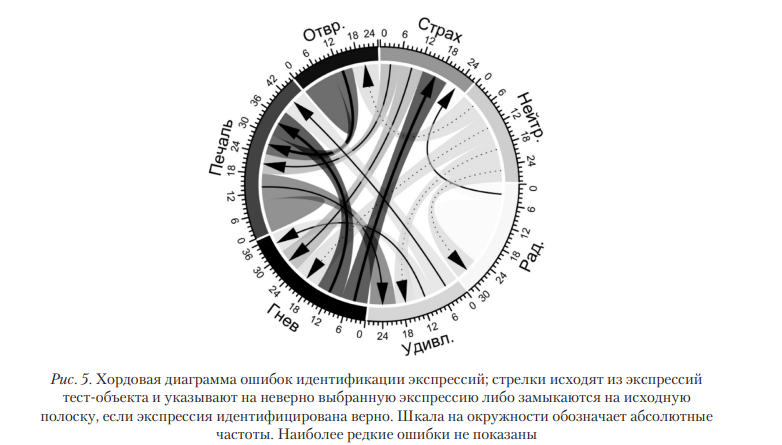
Ошибки идентификации. На рис. 5 представлена структура ошибок идентификации экспрессий в условиях интрасаккадической смены стимула. Из диаграммы можно предположить, что экспрессия радости обладает хорошо различимыми визуальными признаками, поэтому аттрактивность не сказывается на оценках, когда она выступает в качестве альтернативного варианта. Нейтральная экспрессия обладает наименее выраженными визуальными признаками, а при сравнении с другими экспрессиями легко отличима от правильного варианта, почему на нее не указывают другие стрелки на диаграмме. Наиболее часто неверно идентифицируются гнев, удивление и печаль.
Видимые локализации. Поскольку и латеральный стимул, и интрасаккадический объект экспонировались в одной и той же позиции, эффект сжатия визуального пространства отсутствует (рис. 1, фрагменты 2, 3). Существует незначительная тенденция к удлинению видимого расстояния до тест-объекта, но для данного анализа статистической мощности недостаточно. В целом, смена стимула в интрасаккадическом интервале не повлияла на правильность оценки локализации. Повторим, что согласно процедуре эксперимента предмет восприятия (латеральный стимул, длительностью более 100 мс или интрасаккадическое изображение лица, длительностью менее 15 мс), а следовательно и его локализация, произвольно определяются самим наблюдателем.
Обсуждение результатов
Резюмируя итоги проведенных экспериментов, отметим следующее. Во-первых, в амбивалентной ситуации, допускающей произвольную категоризацию последовательно экспонируемых кратковременных выражений лица, первое изображение идентифицируется наблюдателями в 3,5 раза чаще. Учитывая кратное превышение длительности первого (латерально расположенного) изображения над тестовым, этот результат нетрудно отнести к проявлениям прямой зрительной маскировки. Новизна эффекта состоит в том, что маскирующий стимул экспонируется на ближней периферии (±10°) во время исходной фиксации, а тест-объект — в процессе выполнения саккады. В терминах обсуждаемой проблемы это означает, что предсаккадическая информация влияет на интрасаккадическую, резко снижая различительную чувствительность зрительной системы.
Во-вторых, условием проявления маскировки описанного типа является однородность значимого содержания латерального и тестового стимулов. Как мы показали ранее [Barabanschikov, 2019], использование в качестве латерального стимула фиксационного креста (<1°) не влияет радикально на оценку лицевых экспрессий, демонстрируемых во время саккад. Существенно, что введение изображений лица сокращает латентный период вызываемой саккады; появляется индикатор усложнения предметного содержания стимульной ситуации.
В-третьих, хотя частота корректной идентификации тест-объектов находится на уровне ниже шанса, выполненные оценки носят закономерный характер. В ослабленном варианте прослеживаются тенденции, обнаруженные нами в экспериментах, где реактивная саккада запускалась изображением миниатюрного креста: рейтинг распознавания эмоций, структура ошибок, зависимость частоты корректной идентификации от альтернативного варианта ответа, показатели реактивных 10°-саккад (кроме латентного периода) и др. Это означает, что ключевые детерминанты транссаккадического восприятия в рамках нового эксперимента сохраняются, а их совокупный продукт действительно подвержен прямой зрительной маскировке.
Проведенный анализ позволяет вернуться к вопросам природы саккадического подавления. С точки зрения полученных данных, на уровне предметного восприятия значимых событий, падение различительной способности во время саккад не требует апелляции к самостоятельному механизму, ответственному за фрагментацию зрительного процесса. Достаточно сочетания условий, каждое из которых ведет к снижению идентификации лицевых экспрессий. В исследованной ситуации их несколько: 1) сверхкороткая экспозиция тест-объекта; 2) его локализация на периферии; 3) маскирующее влияние латерального изображения лица; 4) сходство лицевых экспрессий на уровне как стимулов, так и ответов. В качестве дополнительных условий выступают: (5) объективное расположение маскера и теста в одной и той же пространственной позиции, а также (6) возможность метаконтраста при появлении прямоугольной рамки для оценки локализации тест-объекта. Каких-либо норм корректности идентификации выражений лица не существует. Это оперативная характеристика, возникающая «здесь и теперь» на основе наличных предпосылок в соответствии с требованиями текущего поведения. В зависимости от складывающихся обстоятельств, уровень и вектор зрительной чувствительности меняется. Например, исключив влияние содержательного маскера, как было сделано нами в ранних экспериментах, можно повысить степень эффективности транссаккадического восприятия, но до определенного уровня, формируемого набором других условий.
Подобная логика реализуется и в исследованиях психофизики транссаккадического восприятия, за исключением одного: главным феноменом здесь выступает саккадическое подавление. Зрительное восприятие сверхкоротких событий зависит от размера стимула, его яркости, локального контраста, соотношения пространственных частот фигуры/фона и их отношений. Существенную роль обратной маскировки (метаконтраста) в саккадическом подавлении продемонстрировал MacKay (1972). Согласно кривым прямой и обратной зрительной маскировки, полученным Crawford (1947), наибольший эффект (1,9—3 лог ед.) наблюдается тогда, когда между стимулом и маскером проходит от 0 до ± 80 мс. Полное отсутствие эффекта маскировки имеет место в стимульном интервале от -10 до -20 мс. Данные получены при угловом размере стимула 0,5°, длительности стимула 10 мс, а маскера — 524 мс (как известно, эффект метаконтраста усиливается, если маскер ярче или имеет более длительную экспозицию). Подобная конфигурация частично воспроизводится в нашем эксперименте: тест-объект экспонируется приблизительно за 10—15 мс до завершения саккады. В нормальных условиях она заканчивается фиксацией нового объекта с возможностью обратной маскировки. Наличие нового объекта позволяет поддерживать стабильность видимого мира и объективирует актуальный центр зрительного поля. При отсутствии объекта фиксации, как в поставленном лабораторном эксперименте, вместо ожидаемой цели воспринимается однородный, «пустой», фон, по отношению к которому информация, полученная интрасаккадически, приобретает статус фигуры и осознается.
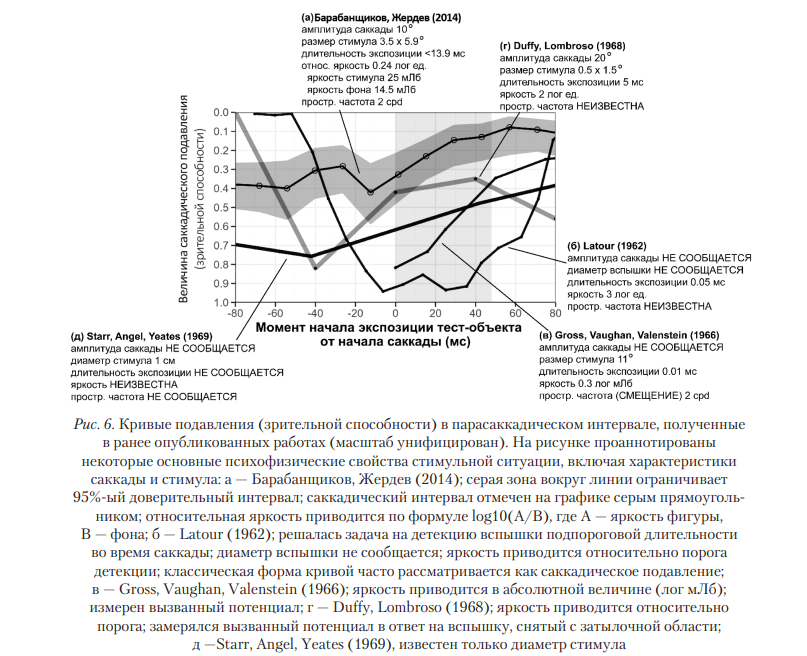
Сравнивая параметры стимулов, использованных другими авторами в сходных экспериментальных ситуациях [Lederberg, 1970; Volkmann, 1962], можно заметить, что яркость стимула в нашей работе [Барабанщиков, 2014] была наименьшей, показав положительную вероятность идентификации. Снизить эффект саккадического подавления удается не только в обсуждаемом эксперименте. Это можно сделать при достижении достаточного углового размера тест-объекта в диапазоне пространственных частот (1,8—3 cpd). При этом длительность экспозиции тест-объекта должна оставаться минимальной и не выходить за его временные границы. Ряд авторов делают вывод, что минимального уровня саккадического подавления можно достичь в условиях предъявления решеток с низкой пространственной частотой (<1 cpd), в условиях же структурированного фона подавление достигает максимума [Mitrani, 1973; Volkmann, 1986; Zimmermann, 2020]. Заметим, что использовавшиеся ранее геометрические примитивы (точки, полосы, вспышки) соответствуют пространственной частоте значимо большей (5—8 cpd [Deubel, 1996; Volkmann, 1962], чем пространственная частота лица в данном угловом размере (2 cpd). На этой пространственной частоте (1,7—2,0 cpd) находится максимум чувствительности к контрасту [Goto, 2001] и цветовым различиям [Okiyama, 2007]. Из работы Burr, Morrone, Ross (1994) следует, что саккадическое подавление наиболее вероятно в диапазоне пространственных частот 0,2—0,8 cpd, тогда как полосу 2,0—10,0 cpd подвергнуть саккадическому подавлению практически невозможно.На рис. 6 приведена динамика зрительной способности («саккадического подавления») до, во время и после начала саккады, полученная в наших работах и в ряде других работ. Видно, что кривая динамики очень непостоянна, а глубина ее спуска, форма и временные границы зависят от психофизических свойств стимула: его яркости, контраста, пространственной частоты, угловых размеров и др. Во всех приведенных работах, кроме Latour, показано снижение значений зрительной способности за 20—40 мс до начала саккады. Напротив, частота верной идентификации для всех стимульных ситуаций растет, приближаясь к максимуму в конце саккады. При экспозиции лицевых экспрессий зарегистрирована более высокая эффективность зрительной способности и более пологая динамика кривой.
Обратим внимание на следующие особенности кривых, представленных на рис. 6.
Во-первых, в большинстве стимульных ситуаций падение чувствительности регистрируется за 20—40 мс до саккады и достигает максимума к ее началу, что совпадает с латентным периодом движений глаз. Во время их перемещения зрительная способность не подавляется, а монотонно растет, восстанавливается. Это означает, что источником подавления (в узком значении термина) является не саккада сама по себе, как иногда считают, а подготовка к ее выполнению, вызванная переносом внимания на новый объект и начавшимся преобразованием зрительного поля. Речь идет о переформатировании воспринимаемого пространства, которое сопровождается ложной локализацией стимула, сжатием пространства в области, прилегающей к новому объекту фиксации, перераспределением четкости и дифференциированности видимых объектов и их отношений, программированием целенаправленной саккады, а также смещением виртуальной позиции наблюдателя. В проведенном нами эксперименте следы «саккадического подавления», которые могли бы проявляться в фактах снижения зрительной способности в зоне исходной фиксации и ослабления прямой зрительной маскировки тестового стимула, не обнаружены.
Во-вторых, радикальные различия в динамике зрительной способности обусловлены спецификой структуры и содержания стимульных ситуаций. Выделяются два уровня организации транссаккадического восприятия, объединенные общей темпоральной схемой: сенсорный и гностический. Первый связан с решением задачи обнаружения стимула (оптических и геометрических примитивов), второй — задачи идентификации, отнесенности стимула к определенной категории. В повседневном поведении обе задачи тесно связаны и перманентно переходят друг в друга.
Таким образом, сложный предсаккадический стимул вызывающий реактивную саккаду, действительно влияет на идентификацию тест-объекта, появляющегося и исчезающего во время саккады, если тест и стимул похожи по содержанию. Зарегистрированный эффект прямой зрительной маскировки исключает «саккадическое подавление», которое ярко проявляется при обнаружении элементарных стимулов. Транссаккадическое восприятие имеет уровневое строение и реализуется на основе общей темпоральной схемы. Зависимость зрительной способности наблюдателя от внешних и внутренних детерминант предоставляет возможность находить сочетания условий, наиболее благоприятные для эффективного восприятия событий в интрасаккадическом диапазоне. Появляется перспектива практического использования зрительной информации, получаемой человеком в ходе выполнения быстрых движений глаз.
Заключение
Совокупность результатов выполненного исследования подтверждает возможность восприятия комплексных экологически валидных объектов во время быстрых движений глаз и характеризует условия, определяющие этот процесс. На материале идентификации эмоциональных экспрессий лица зарегистрировано влияние предсаккадической информации на интрасаккадическую (эффект прямой зрительной маскировки). В вырожденной форме воспроизведены основные тенденции, обнаруженные ранее в сходных экспериментальных ситуациях: рейтинги корректной идентификации модальностей эмоций, структура ошибок, зависимость частоты корректной идентификации от альтернативного выбора ответа, параметры целенаправленных саккад и др. Условием проявления маскировки описанного типа является однородность содержания латерального и тестового стимулов. Использование изображения лица в качестве триггера быстрых движений глаз значимо сокращает латентный период реактивной саккады. Эффект «саккадического подавления» — ключевого феномена в экспериментальных исследованиях обнаружения элементарных оптических свойств и отношений отсутствует. Сравнительный анализ данных, полученных в различных стимульных ситуациях, позволяет ввести представление о сенсорном и гностическом уровнях организации транссаккадического восприятия, объединенных общей темпоральной схемой. Основой динамики зрительной способности до, во время и после выполнения саккады полагается переформатирование воспринимаемого пространства при появлении нового объекта интереса. Саккада не только разделяет и дробит поток зрительной информации на отдельные микрофрагменты, но и одновременно связывает их в монолитное целое. Восприятие — и дискретно, и непрерывно.