«Мониторинг выполнения» (performance monitoring) — широкое понятие, включающее детекцию последствий поведения и инициацию его адаптивных перестроек. Основными электрофизиологическими коррелятами мониторинга выполнения считаются два компонента потенциалов, связанных с событиями (ПСС): ERN (негативная волна, связанная с ошибкой) и Pe (позитивная волна, связанная с ошибкой). ERN — это негативная волна, максимально выраженная в центральных отведениях, пик которой приходится примерно на 100 мс относительно момента начала реакции. Хотя ERN, как правило, следует за ошибками [Falkenstein, 1991; Gehring, 1993], эта негативная волна не является специфичной именно для ошибок: аналогичный компонент ПСС может следовать за правильными ответами [Vidal, 2000], в этом случае он называется CRN (негативность, связанная с правильным ответом). Этот компонент имеет наибольшую выраженность в состоянии неопределенности [Pailing, 2004], при котором система мониторинга выполнения не может корректно произвести оценку правильности выполняемого ответа. Pe представляет собой медленную положительную волну во временном окне от 150 до 350 мс относительно начала реакции [Falkenstein, 1991; Falkenstein, 2000]. Данный компонент ПСС может отражать осознание результата совершенного действия (например, осознание ошибки). Pe можно разделить на два подкомпонента: ранний Pe (150—230 мс) с фронто-центральной локализацией и поздний Pe (230—300 мс), смещенный в сторону затылочной области [van Veen, 2002].
Об осознании совершенной ошибки говорят в том случае, если человек может сообщить об ошибочности действия после его совершения. Как правило, осознанность ошибки определяется при помощи опроса после реализации поведенческого ответа — как, например, в работе Шефферса и Колеса [Scheffers, 2000]. Связь между ERN и осознанием ошибки не вполне ясна. Согласно результатам некоторых исследований, ERN сильнее выражен в случае осознанных ошибок, чем в случае неосознанных [Scheffers, 2000], тогда как в других работах сообщают об отсутствии такой связи [Endrass, 2007]. В отличие от ERN, связь между Pe и осознанием ошибки показана во множестве исследований [Wessel, 2012]. Pe отражает накопление субъективных свидетельств, которые могут привести к осознанию ошибки [Steinhauser, 2010].
Успешность мониторинга выполнения задачи зависит от качества обработки и кодирования стимула [Scheffers, 1999]. Нарушение обработки увеличивает трудность задачи, что приводит к увеличению амплитуды N2 и P3 [Folstein, 2008]. Комплекс N2/P3, по-видимому, аналогичен комплексу ERN/Pe [Falkenstein, 2000]. Компоненты N2 и ERN, по всей видимости, генерируются в одних и тех же зонах мозга (в основном передняя поясная кора) и имеют сходную функциональную роль [Gruendler, 2011]. В рамках теории мониторинга конфликтов считается, что N2 отражает конфликт, предшествующий ответу [Yeung, 2014], а ERN отражает конфликт, следующий за ответом [Danielmeier, 2009].
Очевидно, что процесс идентификации стимула и сопоставления его с правилами задачи необязательно заканчивается в момент принятия решения и может продолжаться даже после того, как моторная программа ответа была запущена. Таким образом, уже после того, как моторный ответ был инициирован или даже осуществлен, может измениться ментальная репрезентация правильного ответа, что, в свою очередь, может повлечь коррекцию или торможение моторной программы — если она приобретает внутренний статус ошибочного действия. Логично предположить, что осознание ошибки и ее исправление могут быть связаны, поскольку оба эти процесса основаны на обработке характеристик поступившего стимула и сопоставлении их с правилами задачи, которое продолжается после начала моторного ответа. Тем не менее, связь ERN и Pe с исправлением ошибочных ответов изучена недостаточно [Kieffaber, 2016].
Исправление ошибок может принимать форму нового, «корректирующего» ответа, который следует за первоначальным неправильным ответом или накладывается на него [Kieffaber, 2016]. Но чаще наблюдается лишь некоторое торможение движения. На поведенческом уровне процесс остановки может проявляться в уменьшенной силе нажатия клавиш в случае ошибок [Rabbitt, 1978], в увеличении времени реакции в ошибочных реализациях или в «постошибочном замедлении» [Novikov, 2015; Novikov, 2017; van Driel, 2012]. Аналогичные процессы при ответах, формально засчитанных как правильные, практически не изучены; однако среди таких ответов могут быть такие, которые привели в конечном итоге к правильному результату, но в процессе инициации и исполнения подвергались дополнительной проверке и/или коррекции системой мониторинга выполнения.
Регистрация нажатий на клавиши, как метод записи ответов испытуемых, предполагает измерение только такого показателя, как время реакции, которого может быть недостаточно для изучения коррекции и/или остановки выполняемого ответа. Регистрация перемещения курсора компьютерной мыши является потенциально более универсальным методом записи ответов по сравнению с регистрацией нажатия клавиш [Freeman, 2011]. Киффабер с соавторами [Kieffaber, 2016] использовали регистрацию координат курсора компьютерной мыши в методике с подсказкой, задающей переключение установки на выполнение задачи (cued task-set switching). Они сообщили, что Pe возникала в случае исправленных ошибок, а в случае совершенных ошибок Pe не наблюдалось; при этом оба типа ошибок вызывали ERN. Торможение выполняемой реакции как таковое электрофизиологически не исследовалось. Таким образом, связь ERN/CRN и Pe с коррекцией/остановкой ответа все еще недостаточно изучена [Kieffaber, 2016]; при этом крайне ограниченными являются сведения о мозговых процессах, предшествующих ответу и связанных с уровнем неопределенности, который, в свою очередь, влияет на последующий мониторинг выполнения.
В данном исследовании мы использовали регистрацию перемещения курсора компьютерной мыши при выполнении испытуемыми слуховой конденсационной задачи, аналогичной той, которая была использована нами в наших предыдущих исследованиях и сопровождалась традиционной регистрацией нажатий на клавиши [Novikov, 2015; Novikov, 2017]. Благодаря регистрации перемещения курсора компьютерной мыши мы получили возможность оценить два поведенческих показателя, связанных с ответом: время начала движения курсора компьютерной мыши и продолжительность перемещения. Мы предположили, что время начала движения аналогично стандартному времени реакции, а продолжительность движения мы рассматривали как дополнительный показатель, чувствительный к торможению ответа после его инициализации.
В настоящем исследовании мы рассмотрели два аспекта мониторинга выполнения: один связан с неопределенностью во время обработки стимула и инициации ответа, т. е. с формированием исходных данных для мониторинга выполнения; другой связан с поведенческими корректировками, которые следуют за первоначальной внутренней оценкой моторной программы после ее запуска. Поскольку конденсационная задача обеспечивает высокий уровень внутренней неопределенности [Novikov, 2017], мы ожидали, что эффекты коррекции/ торможения ответа могут наблюдаться и при правильных ответах.
Мы выдвинули следующие гипотезы.
1. Известно, что состояние неопределенности/конфликта перед ответом приводит к увеличению времени реакции и амплитуды N2 [Chernyshev, 2016; Yeung, 2014], а также вызывает нарушение внутренней детекции ошибок [Novikov, 2017]. Таким образом, мы ожидали, что более позднее время начала ответа будет сопровождаться увеличением амплитуды N2 и снижением амплитуды ERN/CRN.
2. Мы рассматривали увеличение продолжительности движения как следствие торможения уже запущенной реакции, которое может быть связано с осознанием ошибки; таким образом, мы ожидали, что Pe будет сильнее выражен для ответов с увеличенной продолжительностью движения.
Методика
В эксперименте принял участие сорок один здоровый испытуемый (21,6 ± 3,7 года). Все участники были правшами, имели нормальное или скорректированное до нормального зрение, нормальный слух и сообщали об отсутствии слуховых, неврологических и психических расстройств. Эксперименты проводились в соответствии с Хельсинкской декларацией и ее поправками и были одобрены этическим комитетом. Перед экспериментом участники подписывали информированное согласие.
Во время эксперимента испытуемые выполняли модифицированную слуховую конденсационную задачу [Novikov, 2015; Novikov, 2017]. Конденсационная задача построена таким образом, что выбор поведенческого ответа осуществляется испытуемыми на основе сочетания признаков стимулов, а не отдельных признаков; такая организация задачи создает высокую когнитивную нагрузку [Garner, 1974; Gottwald, 1975; Posner, 1964].
Слуховые стимулы предъявляли участникам с помощью программного обеспечения E-Prime (Psychology Software Tools, Inc., США) через внутриканальные наушники. В эксперименте использовали четыре предварительно записанных слуховых стимула, которые представляли собой комбинации двух вариантов высоты («низкий» — «ля» первой октавы, 440 Гц и «высокий» — «до» второй октавы, 523 Гц) и двух вариантов тембра (каллиопа и виолончель). Длительность стимулов составляла 40 мс, время нарастания и спада было равно 10 мс, а уровень звукового давления — 95 дБ. Громкость уравнивали между стимулами по среднеквадратичной амплитуде сигнала.
Испытуемые выбирали ответ, перемещая курсор мыши из исходного положения в левую или правую зону ответа (рис. 1) в соответствии с правилом, отраженным на схеме соответствия «стимул—реакция» (табл.). Границы зон ответа были заданы таким образом, что расстояние между начальной точкой и зонами ответа не зависело от угла перемещения курсора. Курсор мыши был скрыт во всех экспериментальных блоках, кроме первого обучающего блока (см. ниже). Координаты курсора мыши непрерывно регистрировались с пространственным разрешением 800 точек на дюйм и частотой дискретизации 125 Гц при помощи программного обеспечения E-Prime.
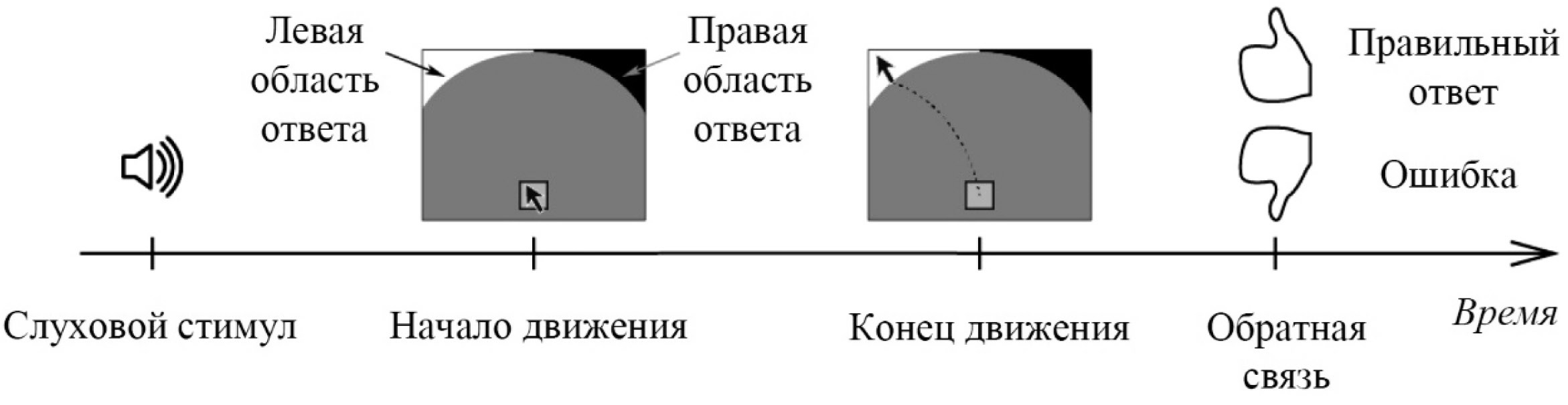
Рис. 1. Схема эксперимента
Таблица
Схема соответствия «стимул-реакция» в экспериментальной задаче
|
Тембр |
Тон |
|
|
Низкий (440 Гц) |
Высокий (523 Гц) |
|
|
Каллиопа |
Левая область ответа |
Правая область ответа |
|
Виолончель |
Правая область ответа |
Левая область ответа |
Инструкция испытуемым не содержала ни требование быстрого ответа, ни требование (или рекомендации) отвечать наугад в случае сомнений. Однако, отсутствие ответа в течение 4000 мс после предъявления стимула засчитывалось как пропуск, и начиналась следующая реализация.
Эксперимент состоял из двух обучающих и шести экспериментальных блоков. Первый обучающий блок включал 15 реализаций; второй обучающий блок и экспериментальные блоки включали по 100 реализаций. Курсор мыши, «стартовая» область и области ответа были представлены на экране только во время первого обучающего блока. Обратную связь испытуемые получали только в обучающих блоках. Если испытуемый в обучающих блоках не достигал критерия обученности (> 60% правильных ответов), то эксперимент прекращали и результаты данного испытуемого исключали из анализа. Во время экспериментальных блоков курсор мыши, «стартовая» область и области ответа были скрыты и обратная связь отсутствовала.
Во время эксперимента регистрировали электроэнцефалограмму (ЭЭГ) с помощью системы NVX-52 (Медицинские компьютерные системы, Россия) с программным обеспечением Neocortex Pro (Нейроботикс, Россия), запись осуществлялась с 40 электродов, расположенных по модифицированной международной системе 10—10. Кроме того, производилась запись с четырех электроокулографических электродов. Референтные электроды находились на мочках уха, заземляющий электрод — в лобной части головы, между электродами Fpz и Fcz. Импеданс был ниже 10 кОм для всех каналов. Регистрация ЭЭГ выполнялась в диапазоне частот 0,5—200 Гц с частотой дискретизации 1000 Гц.
В рамках исследования мы рассматривали два поведенческих результата реализаций: правильные ответы (перемещение курсора в релевантную область ответа) и ошибки (перемещение курсора в нерелевантную область ответа); пропуски ответа были исключены из анализа. Для каждой реализации были рассчитаны два показателя ответа: время начала движения (от момента предъявления стимула до начала перемещения курсора компьютерной мыши) и продолжительность движения (от начала перемещения курсора компьютерной мыши до момента вхождения курсора мыши в любую из областей ответа).
Для каждого испытуемого рассчитывали среднее значение обоих показателей при выполнении экспериментальных блоков — с учетом того, был ли ответ правильным или ошибочным. Для каждого показателя индивидуальные средние значения сравнивались между правильными и ошибочными ответами при помощи парного t-критерия. Кроме того, для каждого испытуемого по совокупности отдельных реализаций (trial-to-trial) мы вычислили корреляцию Пирсона между временем начала движения и продолжительностью движения для правильных ответов, а затем определяли достоверность отличия индивидуальных коэффициентов корреляции от нуля, используя t-критерий для одной выборки. В анализ поведенческих данных вошли 24 испытуемых. Распределение индивидуальных средних значений указанных показателей по выборке испытуемых не отличалось от нормального (тест Колмогорова—Смирнова, p >> 0,05).
В анализ ЭЭГ были включены только данные из экспериментальных блоков. Данные ЭЭГ анализировали с помощью собственных скриптов с использованием внутренних функций программных пакетов EEGLAB [Delorme, 2004] и ERPLAB [Lopez-Calderon, 2014] для среды MATLAB (MathWorks, США). Основные артефакты ЭЭГ были удалены вручную, а прочие артефакты, в том числе электроокулографические и электромиографические, были удалены с помощью анализа независимых компонент (ICA) с использованием собственных скриптов на базе внутренних функций EEGLAB [Delorme, 2004].
Потенциалы, связанные с событиями (ПСС), были вычислены путем когерентного усреднения сигнала ЭЭГ относительно момента предъявления стимула, а также относительно ответа (момента начала движения).
Для каждого испытуемого мы вычислили ПСС для правильных и ошибочных ответов в целом. Кроме того, для каждого испытуемого мы вычисляли ПСС отдельно для следующих условий (только правильные ответы):
— «ранние» (выборка из 25% правильных ответов с наименьшим временем инициализации движения);
— «поздние» (выборка из 25% правильных ответов с наибольшим временем инициализации движения);
— «короткие» (выборка из 25% правильных ответов с наименьшей продолжительностью движения);
— «длительные» (выборка из 25% правильных ответов с наибольшей продолжительностью движения).
Верхние и нижние квартили реализаций по поведенческим параметрам вычисляли после удаления эпох ЭЭГ, содержащих артефакты. Таким образом, для каждого испытуемого было включено в анализ приблизительно равное количество «ранних», «поздних», «коротких» и «длинных» ответов (разница между условиями могла составлять максимум одну реализацию, если общее количество реализаций не являлось кратным 4). Для каждого из условий было включено в анализ 134 ± 25 реализаций (среднее ± стандартное отклонение по группе испытуемых).
Коррекция ПСС производилась относительно фонового уровня, рассчитанного отдельно для каждого условия как среднее значение сигнала в интервале от -100 до 0 мс относительно момента предъявления стимула.
Для анализа ПСС относительно момента предъявления стимула была выбрана область интереса для компонента N2 — Fcz, 230—300 мс (см. данные предыдущего исследования [Chernyshev, 2016]). Для анализа ПСС относительно ответа (начала движения) были выбраны три пространственно-временные области интереса: ERN/CRN — Fcz, 10—110 мс; ранний Pe—Fcz, 120—260 мс; поздний Pe—Pz, 265—430 мс. Выбор областей интереса был основан на предыдущих исследованиях ЭЭГ с регистрацией перемещения курсора компьютерной мыши [Kieffaber, 2016]. Значение амплитуды каждого компонента вычисляли как среднее значение потенциала на соответствующем канале внутри выбранного временного интервала.
Мы сравнили индивидуальные амплитуды компонентов ПСС, усредненные для каждого испытуемого, с помощью парного t-критерия для следующих контрастов: правильные ответы против ошибок, «ранние» правильные ответы против «поздних» правильных ответов, «короткие» правильные ответы против «длительных» правильных ответов. В анализ данных ПСС вошли 16 испытуемых. Распределение индивидуальных средних значений амплитуд компонентов ПСС по выборке испытуемых не отличалось от нормального ни для одного из условий (тест Колмогорова—Смирнова, p >> 0,05).
Результаты
Из поведенческого анализа были исключены данные испытуемых, общая успешность выполнения задачи которых составляла более 95%, а также данные испытуемых, успешность выполнения задания которых составляла менее 50% как минимум на один из четырех стимулов. Таким образом, анализ поведенческих данных был выполнен на выборке из 24 участников. В среднем эти участники совершали 88,0 ± 7,3% правильных ответов и 11,2 ± 6,5% ошибок от общего числа реализаций. Кроме этого, из анализа исключались реализации со временем начала движения менее 300 мс или более 3000 мс с момента предъявления стимула, а также реализации с продолжительностью движения более 1000 мс.
Среднее время начала движения курсора было больше для ошибок по сравнению с правильными ответами (1324 ± 78 мс против 1101 ± 59; t23 = 7,83; р < 0,001); продолжительность движения курсора также была больше для ошибок (217 ± 13 против 197 ± 10 мс; t23 = 4,83; р < 0,001).
Среднее значение индивидуальных коэффициентов корреляции между временем начала движения и продолжительностью движения было равно 0,01 ± 0,1, что не отличалось значимо от нуля (t = 0,632; р > 0,05). Это говорит о том, что время начала движения и продолжительность движения не зависят друг от друга и могут рассматриваться как два отдельных поведенческих показателя.
Лишь менее 1% реализаций в данном исследовании соответствовали критерию исправленных ошибок, предложенного Киффабером с соавторами [Kieffaber, 2016] и основанного на детекции перелома направления траектории движения курсора мыши, поэтому в дальнейшем анализе мы не выделяли исправленные ошибки в отдельный тип ответов.
Далее мы исключили данные 8 испытуемых из анализа ПСС в связи с артефактами в записи ЭЭГ. Таким образом, анализ данных ПСС был проведен на выборке из 16 участников.
Различий в амплитуде N2 между правильными и ошибочными ответами не было выявлено (p > 0,05) (рис. 2А). Однако компонент N2 был значительно сильнее выражен для «поздних» правильных ответов по сравнению с «ранними» правильными ответами (t15 = 7,33; p < 0,001) (рис. 2Б). На топографических картах заметна фронто-центральная негативность для «поздних» ответов, но не для «ранних» (рис. 3А). Не было выявлено различий между «короткими» и «длительными» правильными ответами (p > 0,05) (рис. 2В).
Различий в амплитуде компонентов CRN и ERN при правильных и ошибочных ответах соответственно (рис. 2Г) обнаружено не было (р > 0,05). CRN был сильнее выражен в случае «ранних» правильных ответов по сравнению с «поздними» правильными ответами (t15 = 3,32; p = 0,004) (рис. 2Д); на топографических картах видна фронто-центральная негативность в интервале CRN при обоих условиях, но имеющая большую выраженность для «ранних», чем для «поздних» ответов (рис. 3Б). Различий по амплитуде CRN при сравнении «коротких» и «длительных» ответов выявлено не было (р > 0,05) (рис. 2Е).
Сигнал ПСС обнаруживает более положительную динамику во временном интервале раннего Pe для «длительных» правильных ответов по сравнению с «короткими» правильными ответами (t15 = 5,01; р < 0,001) (рис. 2Е, 3В). Во временном окне позднего Pe (270— 425 мс) таких различий не было обнаружено (р > 0,05). Ни для раннего Pe, ни для позднего Pe не было обнаружено различий между правильными ответами и ошибками (p > 0,05) (рис. 2Г); также не было выявлено различий во временных интервалах раннего и позднего Pe между «ранними» и «поздними» правильными ответами (рис. 2Д).
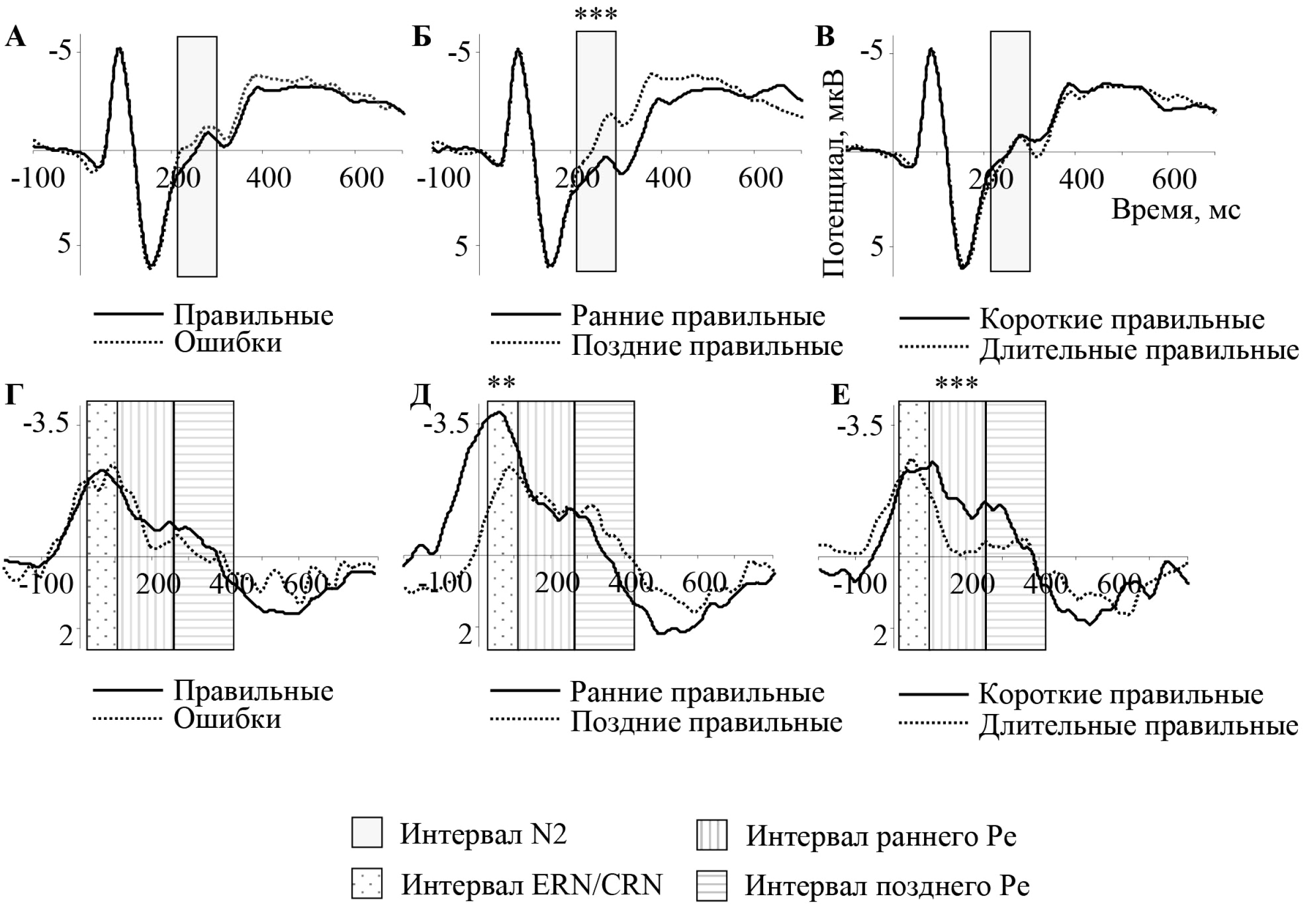
Рис. 2. Потенциалы, связанные с событиями (ПСС), отведение Fcz
Вверху (А, Б, В) — ПСС, усредненные относительно стимула; затемненным прямоугольником обозначен временной интервал N2.
Внизу (Г, Д, Е) — ПСС, усредненные относительно реакции; прямоугольниками обозначены
временны: е интервалы ERN/CRN, ранний Pe и поздний Pe (слева направо).
А, Г — сравнение правильных ответов против ошибочных ответов.
Б, Д — сравнение «ранних» правильных ответов против «поздних» правильных ответов.
В, Е — сравнение «коротких» правильных ответов против «длительных» правильных ответов.
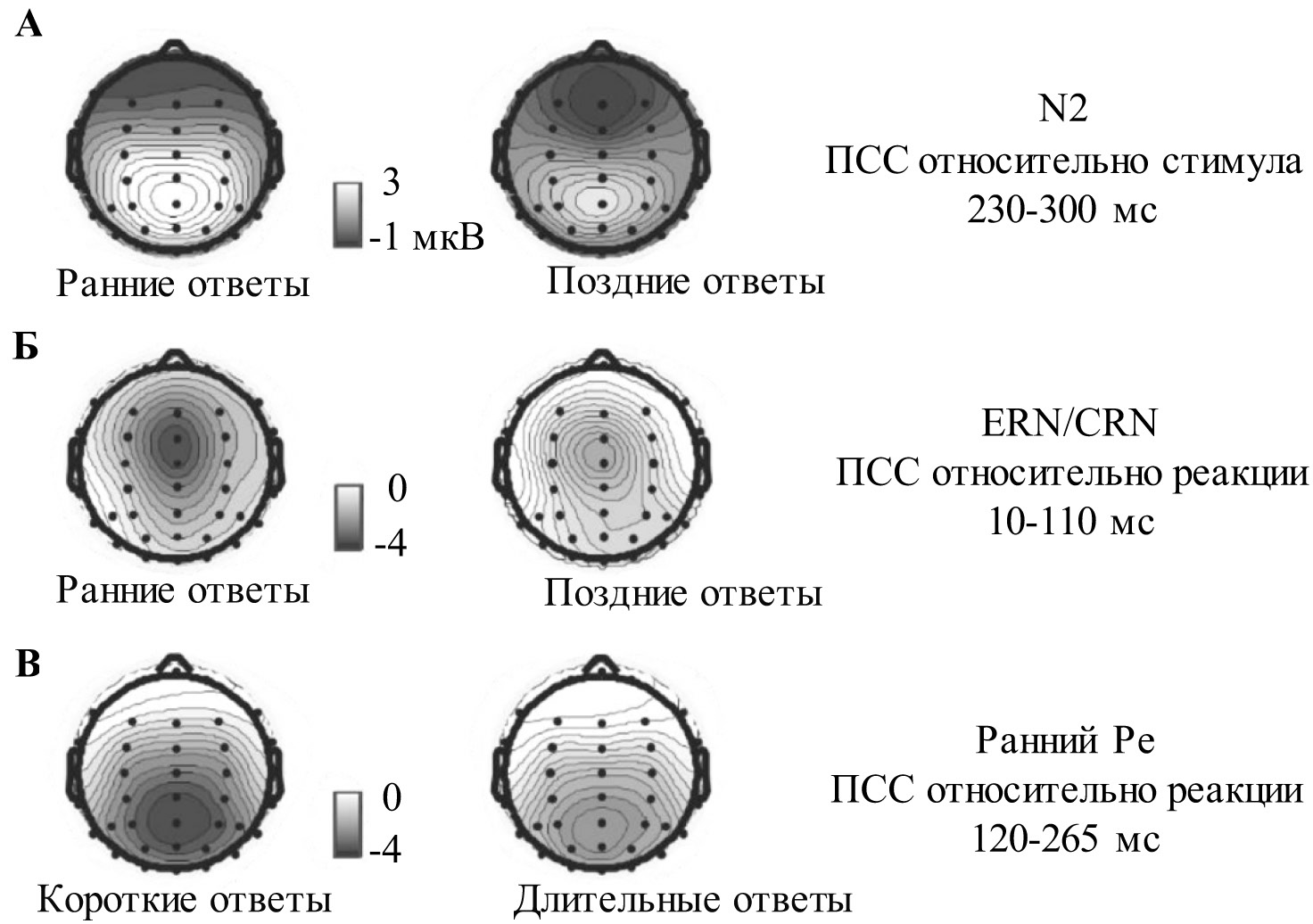
Рис. 3. Топографические карты (средняя амплитуда сигнала во временном интервале компонента): А — N2 для «ранних» (слева) и «поздних» (справа) правильных ответов;
Б — CRN для «ранних» (слева) и «поздних» (справа) правильных ответов;
В — ранний Pe для «коротких» (слева) и «длительных» (справа) правильных ответов
Обсуждение результатов
В настоящем исследовании мы использовали регистрацию перемещения компьютерной мыши для записи ответов испытуемых во время выполнения слуховой конденсационной задачи — идентичной той, которая использовалась в наших предыдущих исследованиях [Novikov, 2015; Novikov, 2017]. Регистрация перемещения курсора компьютерной мыши позволила нам зарегистрировать два поведенческих показателя для каждой реализации: время начала движения и продолжительность движения. Значения времени начала движения согласуются со значениями стандартного времени реакции (при нажатии клавиш), полученными в наших предыдущих исследованиях, в которых также использовалась слуховая конденсационная задача [там же]. Таким образом, подтвердилось выдвинутое нами предположение о том, что время начала движения мыши можно интерпретировать как меру, аналогичную обычному времени реакции при решении когнитивной задачи. Кроме того, можно сделать вывод, что продолжительность движения является отдельным независимым показателем, не связанным со временем начала движения. Оба показателя были выше для ошибок по сравнению с правильными ответами.
Формы сигнала компонентов ERN и CRN были хороню выражены (рис. 2); статистически значимых отличий между правильными и ошибочными реализациями не наблюдалось. Известно, что значения амплитуды компонентов ERN и CRN приблизительно равны в условиях высокой неопределенности [Chernyshev, 2016]. Таким образом, выполнение слуховой конденсационной задачи характеризуется достаточно высоким уровнем неопределенности, как и предполагалось ранее [Novikov, 2015].
Наблюдалось увеличение амплитуды N2 одновременно со снижением амплитуды CRN в случае «поздних» правильных ответов по сравнению с «ранними» правильными. Эти результаты согласуются с данными Данильмейер с соавторами [Danielmeier, 2009], которые сообщили об отрицательной корреляции между амплитудами N2 и ERN/CRN при выполнении фланговой задачи Эриксена.
Система внутренней детекции ошибок оценивает степень несоответствия между репрезентациями вероятного правильного ответа и фактически выполненного ответа; на электрофизиологическом уровне это несоответствие проявляется как ERN/CRN. Предположительно, в условиях высокой неопределенности возникает более сильный конфликт перед ответом, который проявляется в увеличении N2. В данных условиях не только увеличивается время реакции, но и ухудшается репрезентация правильного ответа, что нарушает работу внутренней детекции ошибки, что, в свою очередь, уменьшает амплитуду ERN/CRN. В условиях низкой неопределенности будет наблюдаться обратная зависимость.
Форма сигнала ПСС характеризовалась более выраженной положительной динамикой во временном интервале раннего Pe для «длительных» ответов по сравнению с «короткими». Соответственно, мы предполагаем, что ранний Pe связан с торможением (остановкой) выполняемого ответа; этот результат согласуется с экспериментальными данными, связывающими Pe с осознанием ошибки [Wessel, 2012]. В настоящем исследовании данный эффект наблюдался при положительных исходах действия и, вероятно, отражал пересмотр/коррекцию действия, в сочетании с осознанием потенциальной вероятности совершения ошибки.
Выводы
Итак, применение метода регистрации перемещения курсора компьютерной мыши позволило измерить два взаимодополняющих независимых поведенческих показателя: время начала движения («ранние» и «поздние» ответы) и продолжительность движения («короткие» и «длительные» ответы). N2 был увеличен, а CRN снижен для «поздних» правильных ответов по сравнению с «ранними» правильными; данный результат можно объяснить тем, нто «поздние» ответы характеризуются более высоким конфликтом между альтернативами решений перед ответом и более высокой неопределенностью, в то время как для «ранних» ответов характерен более низкий уровень неопределенности. Ранний Pe был сильнее выражен для «длительных» ответов по сравнению с «короткими», этот эффект может объясняться вовлечением механизмов подавления выполняемого ответа вследствие вовлечения механизмов мониторинга выполнения.