Введение
Практика поэтапного формирования навыков широко распространена и часто используется в экспериментальной психологии. Как правило, поэтапное обучение характеризуется постепенными переходами от простого к сложному. В этих случаях изначально на первых этапах усвоенные правила оказываются уже неэффективными или недействующими для последующих этапов. Альтернативой такому многоэтапному формированию какого-либо навыка является обучение одноэтапное, когда вся проблемная ситуация представлена целиком с самого начала и изменения правил от этапа к этапу не происходит. Являются ли идентичными мозговые механизмы приобретения навыков этими двумя способами?
Мозговым субстратом приобретения какого-либо навыка является совокупность нейронов, изменяющих свои свойства, в том числе электрические. В соответствии с системноселекционной теорией обучения (Швырков, 1995), формирование нового поведения при научении включает в себя формирование новой функциональной системы – системогенез; на нейронном уровне этому процессу соответствует формирование специализаций нейронов относительно этой системы. Специализация нейронов проявляется в возникновении у ранее молчащих нейронов активаций, неизменно появляющихся при осуществлении сформированного поведенческого акта. В настоящее время известно большое число примеров поведенческих специализаций нейронов. Это нейроны «восприятия определенных персонажей» (Quiroga et al., 2007; Gelbard-Sagiv et al., 2008) у людей, актов «социального контакта» с определенными особями в стаде у обезьян (Perrett et al., 1996), актов инструментального поведения у животных (Александров и др., 1997), актов ухода за новорожденными ягнятами у овец (Kendrick et al., 1992), актов реализации концепта «гнезда» (Lin et al., 2007) и другие.
Однако можно предположить, что научение включает в себя не только формирование новых специализаций у нейронов, но и реорганизацию уже существующих нейронных групп. Нейронные механизмы обучения, таким образом, могут предполагать наличие двух видов процессов: формирование новых специализаций нейронов и аккомодационную реконсолидацию нейронов, уже имеющих системную специализацию (Alexandrov et al., 2001). Поскольку процессы аккомодационной реконсолидации затрагивают нейроны, специализированные относительно приобретенного ранее опыта, при доучивании или переучивании именно в процессах аккомодационной реконсолидации должны проявляться различия в мозговых субстратах, сформированных поэтапно или одноэтапно.
Механизмы аккомодационной реконсолидации окончательно не изучены. Однако есть основания полагать, что одним из таких механизмов может быть изменение экспрессии генов в нейронах (Svarnik et al., 2008; Сварник, 2010). Изменение экспрессии генов в нейронах может быть детектировано по экспрессии непосредственного раннего гена c-fos, продукт активности которого – белок Fos – является транскрипционным фактором, индуцирующим запуск экспрессии многих генов (Анохин, 1997). Целью данной работы было получение ответа на вопрос о том, тождественны ли мозговые механизмы приобретения навыков одно- и многоэтапным способами, посредством выявления связи между характеристиками реорганизации предыдущего опыта (оцененного по нейрогенетическим изменениям) при переучивании и числом этапов предварительного обучения. Для того чтобы получить ответ на сформулированный выше вопрос, мы обучали животных двух групп одному и тому же поведению разными способами – в один или несколько этапов, а затем переучивали животных обеих групп сходному навыку и оценивали число нейронов, подвергающихся изменению экспрессии генов.
Метод
Животные. В работе использовались капюшонные крысы Long-Evans массой 200– 300 грамм. На время проведения эксперимента животных помещали в индивидуальные клетки размером 40х25х20 см. Обучение проводилось после периода адаптации. С момента начала обучения животные экспериментальных групп находились на пищевой депривации. Потеря веса за период обучения не превысила 20 %. Животные контрольной группы находились в домашних клетках вивария в течение всего экспериментального периода и имели свободный доступ к пище и воде.
Экспериментальная клетка. Экспериментальная клетка содержала две автоматические кормушки, расположенные в углах передней стенки клетки, и две педали, находящиеся в противоположных углах дальней стенки клетки. Вне клетки находилась кнопка экспериментатора, нажатие которой позволяло подавать кормушку с пищей в любое время. Поведение животных регистрировалось при помощи цифровой видеокамеры. Анализ видеозаписи поведения был осуществлен с использованием программы Easy Track Light 1.0.3.
Обучение. Животные предварительно обучались пищедобывательному поведению нажатия на педаль на одной стороне экспериментальной клетки. Животные первой экспериментальной группы обучались данному навыку многоэтапно (группа «5 этапов») в течение пяти дней: во время первой 30-минутной экспериментальной сессии (первый день) животные получали пищу за нахождение рядом с кормушкой, во второй день – за поворот головы в сторону педали, в третий – за отход от кормушки, на четвертый день – за подход к педали, затем, на пятый, – за нажатие на педаль. Животные второй экспериментальной группы обучались данному навыку одноэтапно (группа «1 этап»). Одноэтапное обучение также проводилось в течение пяти дней, но на протяжении всего этого времени животные получали пищу лишь в случае нажатия на педаль. После приобретения данного навыка животные обеих групп тренировались в выполнении этого пищедобывательного поведения в течение последующих пяти дней.
После стабилизации навыка на первой стороне экспериментальной клетки животные должны были обучиться аналогичному навыку на второй стороне клетки (с использованием противоположных педали и кормушки) в течение последней 30-минутной экспериментальной сессии. При этом педаль и кормушка на первой стороне экспериментальной клетки не функционировали.
Иммуногистохимия. Для исследования экспрессии раннего гена c-fos после последней экспериментальной сессии крысы были помещены в домашние клетки на 1 ч 15 мин, после чего усыплены ингаляционным эфирным наркозом и декапитированы. Непосредственно после этого мозг животных был извлечен и заморожен в жидком азоте. Животные контрольной группы (группа «контроль») были взяты из домашней клетки непосредственно перед декапитацией.
Экспрессию гена c-fos оценивали в ретросплениальной коре по наличию продукта его экспрессии – белку c-Fos. Ретросплениальная кора была выбрана для анализа, поскольку эта область коры головного мозга крыс характеризуется относительно большим числом нейронов, электрическая активность которых специфически связана с выполнением данного пищедобывательного навыка (Svarnik et al., 2005).
Были приготовлены фронтальные криостатные срезы ретросплениальной коры головного мозга крыс толщиной 20 мкм. Взятые срезы были расположены на уровне –4.5 от брегмы. После фиксации в 4-процентном параформальдегиде и промывки срезы помещались в установку для иммуногистохимической реакции (Sequenza Immunostinding Center, Shandon, UK) и прединкубировались в фосфатном буфере с добавлением 2,5-процентной нормальной козьей сыворотки для снижения неспецифического окрашивания. Инкубация с первичными антителами c-Fos (АВ-5, oncogene Science, USA) в разведении 1:2000, по 95 мкл на стекло, проводилась в течение 18–20 часов при комнатной температуре. После инкубации с первичными антителами срезы были промыты и инкубированы со вторичными (Vectastain Elite ABC KIT, Vektor, USA) в разведении 1:200, по 95 мкл на стекло. Инкубация проводилась в течение двух часов. После промывки стекла со срезами были инкубированы со стрептавидинбиотиновым комплексом (Vectastain Elite ABC KIT, Vektor, USA), после чего помещены в штативы для выявления пероксидазной реакции диаминобензидином (Sigma, USA). После появления окраски (7 мин) стекла были дегидратидированы проведением через серию спиртов восходящей концентрации и ксилол, а затем заключены под покровные стекла.
Срезы оцифровывались при 10-кратном увеличении на микроскопе Olympus BX-50 с помощью высокоразрешающей CCD камеры (Nikon DMX-1200) и вводились в компьютер для анализа распределения иммунопозитивных клеток в мозге. Окрашенные клетки в исследуемых областях мозга были подсчитаны на компьютере с помощью морфометрической программы Image Pro 3.0 (Media Cybernetics Inc., USA).
Для оценки статистической достоверности различий поведенческих показателей и процентов Fos-положительных нейронов в ретросплениальной коре у животных разных групп использовался непараметрический критерий Манна-Уитни. Различия считались достоверными при p<0,05.
Результаты
Поведение животных при обучении на первой стороне экспериментальной клетки. Животные группы «1 этап» и животные группы «5 этапов» предварительно обучились пищедобывательному поведению нажатия на педаль на первой стороне клетки (см. раздел «Метод»). Обучение и в одном, и в другом случае заняло пять дней. На пятый день животные выполняли циклическое пищедобывательное поведение нажатия на педаль. Следующие пять дней животные осуществляли выученное поведение, постепенно увеличивая число нажатий на педаль. Различий в выполнении данного поведения на десятый день между животными обеих экспериментальных групп обнаружено не было.
Поведение животных при обучении на второй стороне экспериментальной клетки. После стабилизации навыка на первой стороне экспериментальной клетки животные обучались этому же пищедобывательному навыку на второй стороне клетки в течение последней 30-минутной сессии. Поведение животных обеих экспериментальных групп было проанализировано при помощи программы Easy Track. Треки поведения репрезентативных животных группы «5 этапов» и группы «1 этап» представлены на рис. 1.
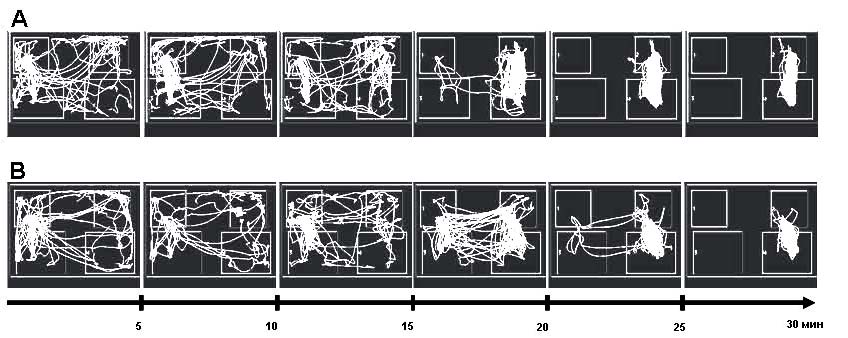
Рис. 1. Пятиминутные треки поведения репрезентативных животных группы «5 этапов» (А) и группы «1 этап» (В) при обучении пищедобывательному навыку на второй (расположенной справа) стороне клетки в течение последней 30-минутной сессии. Верхние квадраты – зоны кормушек, нижние квадраты – зоны педалей
Было обнаружено, что длина треков, т. е. длина пройденного пути за все время поведенческой сессии, у животных группы «1 этап» составляет 15832,8±4065 см (здесь и далее – среднее ± ст. отклонение), а у животных группы «5 этапов» – 10883,6±2026 см. Таким образом, длина трека у животных группы «1 этап» оказалась достоверно больше, чем у животных группы «5 этапов» (z=-1,98; p=0,047) (рис. 2).
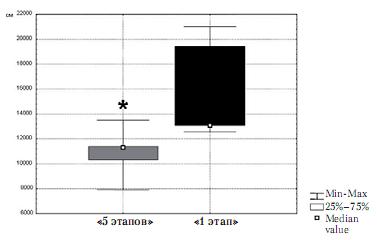
Рис. 2. Длина треков животных группы «5 этапов» и группы «1 этап»
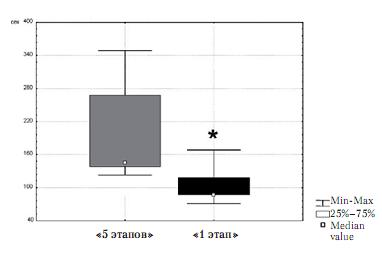
Рис. 3. Суммарное время, проведенное в зоне неэффективной кормушки, у животных группы «5 этапов» и группы «1 этап»
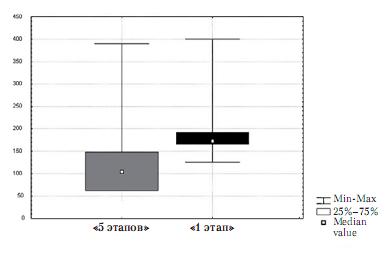
Рис. 4. Число входов в зону эффективной педали у животных группы «5 этапов» и группы «1 этап»
Средняя и максимальная скорости животных в двух экспериментальных группах не различались (z=-1,36; p>0,05).
Поведение животных в течение последней поведенческой сессии было разделено на неэффективное (выполнение ранее выученных поведенческих актов) и эффективное (приобретенное в течение последней сессии). Поведение животных было оценено по четырем категориям: поведение у неэффективной кормушки, поведение у неэффективной педали, поведение у эффективной кормушки и поведение у эффективной педали. Животные обеих групп не различались по числу входов в зону неэффективной педали (z=-1,15; p>0,05) и по суммарному времени, проведенному в зоне неэффективной педали (z=0,73; p>0,05). Животные обеих групп также не различались по числу входов в зону неэффективной кормушки (z=0,10; p>0,05). Однако суммарное время, проведенное в зоне неэффективной кормушки, у животных группы «5 этапов» было достоверно больше, чем у животных группы «1 этап» (z=1,98; p=0,047) (рис. 3).
Животные обеих групп не различались ни по числу входов в зону эффективной кормушки (z=-1,36; p>0,05), ни по времени, проведенному в зоне эффективной кормушки (z=-0,73; p>0,05). Животные обеих групп также не продемонстрировали разницы во времени, проведенном в зоне эффективной педали (z=-0,52; p>0,05), или в числе входов в зону эффективной педали (z=-1,57; p>0,05) (рис. 4).
Нейрогенетические изменения при обучении навыку нажатия на второй стороне экспериментальной клетки. Анализ нейрогенетических изменений проводил-ся на основании наличия продукта экспрессии непосредственного раннего гена c-fos – бел-ка Fos, являющегося маркером изменения экспрессии генов (Анохин, 1997). И у живот-ных группы «5 этапов», и у животных группы «1 этап» число Fos-положительных нейронов в ретросплениальной коре было достоверно выше, чем у контрольных животных, взятых из домашней клетки и не проходивших обучение (z=2,24; p=0,03). Число нейронов, содержащих белок Fos, в ретросплениальной коре у животных группы
«1 этап» оказалось равным 181 ± 35 клеток в 1 мм2, а у животных группы «5 этапов» – 90 ±3 6 клеток в 1 мм2. Число Fos-положительных нейронов у животных группы «1 этап» оказалось достоверно выше, чем у животных группы «5 этапов» (z=2,40; p=0,015) (рис. 5).
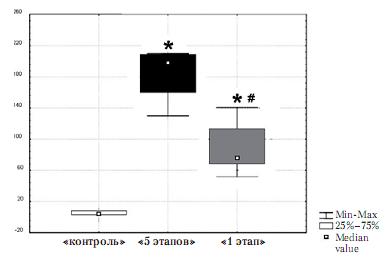
Рис. 5. Число Fos-положительных нейронов в 1 мм2 в ретросплениальной коре у животных контрольной группы, группы «1 этап» и группы «5 этапов». * - p<0.05 по сравнению с группой «контроль», # - p<0.05 по сравнению с группой «1 этап»
Обсуждение
В данном исследовании было обнаружено, что незначительные различия между поведением животных двух экспериментальных групп при переучивании нажатию на педаль на второй стороне экспериментальной клетки сопровождаются достоверными различиями в числе нейронов, содержащих белок Fos и, следовательно, претерпевающих изменения экспрессии генов. Две данные экспериментальные группы различались по предварительной истории формирования навыка нажатия на педаль: одни сформировали навык на первой стороне в один этап, а другие – за несколько этапов.
Результаты, полученные в данном исследовании, свидетельствуют в пользу наличия процессов аккомодационной реконсолидации, т. е. процессов реорганизации уже существующего индивидуального опыта, затрагивающих нейроны, специализированные относительно ранее сформированного поведения. Механизмами, лежащими в основе таких реорганизаций, могут являться не только перестройки экспрессии генов в нейронах, но и структурные изменения в связях между нейронами, например, в числе и паттернах распределения мест контактов между нейронами. Так, например, было показано, что обучение дополнительному навыку не устраняет уже существующие синапсы, а добавляет новые у тех же нейронов (Xu et al., 2009; Yang et al., 2009).
Некоторые параметры поведения животных при формировании навыка на второй стороне клетки в нашем исследовании различались. Было, например, обнаружено, что общая длина побежек, совершенных животными за все время последней экспериментальной сессии, была больше у животных группы «1 этап». Поскольку у животных данных групп не различались ни выраженность неэффективного поведения, ни выраженность эффективного поведения, наличие большей общей моторной активности может свидетельствовать о том, что ориентировочно-исследовательское, или пробное, поведение было у них выражено в большей степени. Для изучения этого вопроса необходимо использовать иные методы анализа поведения. Однако ранее нами было показано, что именно пробное поведение коррелирует с большим числом нейронов, маркированных по белку Fos при научении (Сварник и др., 2007). Кроме того, в работах других авторов многократно было продемонстрировано, что увеличение числа нейронов, содержащих белок Fos, не коррелирует с количеством моторной активности у животных (Kleim et al., 1996; Anokhin, Rose, 1991).
В то же время было обнаружено, что поведение животных группы «5 этапов», по сравнению с животными группы «1 этап», характеризуется увеличением времени, проведенного у неэффективной кормушки. Данный параметр поведения отражает предыдущую историю формирования навыка, поскольку животные группы «5 этапов» многократно проходили через формирование и рассогласование при актуализации данного элемента индивидуального опыта. Каждый пройденный этап формирования этого пищедобывательного поведения характеризовался изменяющимися условиями подачи первой кормушки. Сначала животные обучались тому, что для получения пищи достаточно находиться около кормушки, но затем это правило отменялось, и для получения пищи нужно было отходить от кормушки. После закрепления этого поведения правило снова менялось, и для получения пищи было необходимо подходить к середине стенки клетки, и так далее. Таким образом, предыдущая история формирования данного поведения отражается в параметрах текущего поведения и влияет на состав пробных актов, тестируемых при переучивании на второй стороне экспериментальной клетки.
Проведенные нами ранее эксперименты показали, что число нейронов ретросплениальной коры, специализированных относительно актов пищедобывательного поведения, сформированного одно- и многоэтапным способами, различается: их достоверно больше во втором случае (см.: Alexandrov, 2008). Сопоставление этих и полученных в настоящем исследовании нейрогенетических данных дает основание предположить, что уровень экспрессии ранних генов связан скорее (или в относительно большей степени) с тем, каков объем реорганизации ранее сформированного опыта необходим при новом обучении, чем с тем, каково число нейронов, активация которых этот опыт обеспечивает. Возможно, после ряда пройденных реорганизаций («5 этапов») следующая осуществляется, во всяком случае по изучаемому критерию, с меньшей затратой адаптационных возможностей.
Мы также продемонстрировали, что одноэтапность или многоэтапность предыдущей истории формирования данного навыка не влияют на скорость приобретения навыка на второй стороне клетки. Как в одной, так и во второй экспериментальной группе встречались животные, продемонстрировавшие небольшое число эффективных поведенческих актов, т. е. ненаучившиеся. Однако было бы интересно оценить, насколько прочно сохраняется данный навык в зависимости от того, одноэтапно или многоэтапно он был сформирован с самого начала. Поскольку экспрессия гена c-fos в нейронах является необходимым условием формирования долговременной памяти о навыке (Mileusnic et al., 1996; Grimm et al., 1997; Tolliver et al., 2000), найденные нами различия по экспрессии c-fos у животных двух экспериментальных групп могут свидетельствовать о различной прочности памяти о данном навыке. Этот вопрос требует дальнейшего исследования.