Введение новых объектов в область психологических исследований имеет большое значение для получения знаний об эволюционном генезе и разнообразии психических явлений. Такими новыми объектами могут быть микроорганизмы, начиная с прокариот. Микроорганизмы в зависимости от уровней рассмотрения представляют собой удобную модель, допускающую экспериментирование с использованием инвазивных и деструктивных методик, — в частности, регистрацию таких объективных показателей, как полевые потенциалы [Греченко, 2017; Новик, 1995; Олескин, 2009; Fries, 2015; Masi, 2015].
Микроорганизмы образуют сообщества, одной из форм существования которых являются биопленки. Образование биопленок происходит под влиянием сигналов из окружающей среды и межклеточных взаимодействий [Liu, 2015; Masi, 2015]. Сложность структуры биопленок и многообразие отношений составляющих их микробов превращают их в аналог многоклеточного организма [Шапиро, 1988]. В пользу такого представления свидетельствует также обнаруженное у бактерий известное ранее только у многоклеточных организмов явление апоптоза, задействованное в процессах дифференциации и морфогенеза и контролирующее расположение клеток и создание органов сложной формы [Николаев, 2007; Oleskin, 2019].
При социальном образе жизни в биологических сообществах, независимо от места объектов на эволюционной ступени, отношения между его членами предполагают взаимодействия в форме кооперации, конкуренции и разделения труда [Олескин, 2009; Харитонов, 2014; Шапиро, 1988; Ben-Jacob, 1998; Shapiro, 1995]. Кооперативные связи обеспечивают преимущества в репродукции, получении питания и др. и поэтому широко представлены в биологическом мире на всех уровнях организации живых существ — от генов до социума [Dumas, 2010; Fries, 2015; Liu, 2015; Velicer, 2009]. Кооперация увеличивает общую выгоду популяции благодаря таким процессам, как разделение труда и производство общего блага [Ben-Jacob, 1998; Oleskin, 2019], но в cooбществе индивидуумы могут также конкурировать друг с другом за ограниченные ресурсы, за место в структуре биопленки и др. [Czaran, 2009; Fiegna, 2005]. Наблюдаемые явления связаны с формированием социальных приоритетов, часто входящих в противоречие с необходимостью индивидуального выживания. К кооперативному «альтруистическому» поведению способны и строящие биопленки микроорганизмы, которые по своей организации являются прообразами многоклеточных эукариотных организмов. Мы предполагаем, что разные стадии социальной жизни цианобактерий характеризуются «специальным» типом электрических осцилляций, выраженным определенной частотой и пространственно-временной структурой [Олескин, 2009]. По-видимому, эти параметры полевых потенциалов определяют уровень активности членов сообщества, вовлекаемых в процесс формирования биопленки [Греченко, 2015].
Объектом экспериментов были цианобактерии — одна из древнейших форм жизни на Земле, эволюционный возраст которых, по некоторым данным, приближается к 3,7 млрд лет [Nutman, 2016]. Цианобактерии Oscillatoria terebriformis строят биопленку, отвечающую внешним обстоятельствам и внутрисоциальным потребностям своего сообщества, преобразуя пространственную форму его организации и дифференцируя органоподобные образования [Сумина, 2006; Харитонов, 2014]. Весьма вероятно, что и древнейшие объединения цианобактерий образовывали сложные системы, управлявшие собственным морфогенезом, что позволяло им синхронизированно осуществлять целенаправленное коллективное поведение, в котором пространственные перемещения отдельных нитей определялись целями сообщества.
Задача проведенных нами опытов состояла в выяснении роли уровня синхронизации электрической активности цианобактерий Oscillatoria terebriformis на разных этапах восстановления биопленки.
Методика
Процедура исследования электрической активности. В экспериментах применялся метод регистрации электрической активности с использованием стеклянных электродов, заполненных 1 М KCl. Использовалось от одного до трех электродов, введенных в разные локусы биопленки, что определялось задачами экспериментов. Цианобактерии Oscillatoria terebriformis находились в среде следующего состава (в граммах на литр): NaHCO3 — 3; Na2CO3 — 17; K2HPO4 — 0,5; NaCl — 30; KNO3—2,5; MgSO4 — 0,2; CaCl2 — 0,04; FeSO4 — 0,01, — что наиболее близко к естественному источнику, где была отобрана проба для исследования.
Статистический анализ. Фрагменты записи электрической активности оцифровывались и подвергались спектральному анализу в среде статистической обработки R 3,0 [Греченко, 2019; Shapiro, 1995]. Спектральный анализ выполнялся для исходной записи путем построения периодограммы с использованием быстрого преобразования Фурье [Shumway, 2011]. Для анализа взаимодействий объектов определялись коэффициенты кросскорреляции и когерентности. В работе представлены индивидуальные спектрограммы оцифрованных фрагментов. Длительность оцифрованных участков составляла 3 с. Всего обработано 120 фрагментов записей биопленки Oscillatoria terebriformis, из них 53, полученных при регистрации электрической активности одним электродом, 67 — парой электродов при локализации в актив пых и спокойных областях биопленки.
Результаты
Биопленка, которую формируют цианобактерии, не имеет однотонной окраски: ее цвет варьирует от светло-зеленого до почти бурого и зависит от состояния и активности входящих в ее состав цианобактериальных нитей. Были проведены опыты, в которых при помощи стеклянных электродов из различных областей биопленки Oscillatoria terebriformis, неповрежденной и формируемой в зоне повреждения, регистрировали полевые потенциалы. Получены данные о состоянии осцилляторов в отличающихся по окраске локусах биопленки — активных, имеющих интенсивную зеленую окраску; спокойных (желтовато-зеленых); светло-зеленых, в которых формирование структур только начиналось.
Результаты показали, что частота и амплитуда электрических осцилляций зависит от расположения регистрирующих электродов в локусах повышенной и сниженной активности (в наших опытах в локусах, отличающихся по окраске) (рис. 1, 2А). Электрическая активность представлена колебаниями, частота которых от 0,5 до 45 Гц. Для спокойных областей типичны полевые потенциалы частотой 2—7 Гц, а для активных 20—30 Гц; но возможно как расширение, так и сужение частотного диапазона колебаний электрических потенциалов. Осцилляции могут образовывать паттерны различной длины и иметь разную временную структуру (рис. 1А, 1Б). Для выявления временной структуры веретен применялся автокорреляционный анализ.
Для изучения взаимодействия между различными частями биопленки были выполнены опыты по регистрации потенциалов одновременно из двух локусов. Наличие функциональной связи между разными зонами биопленки выявлялось при помощи коэффициентов кросскорреляции, затем эти связи подвергались и когерентному анализу.
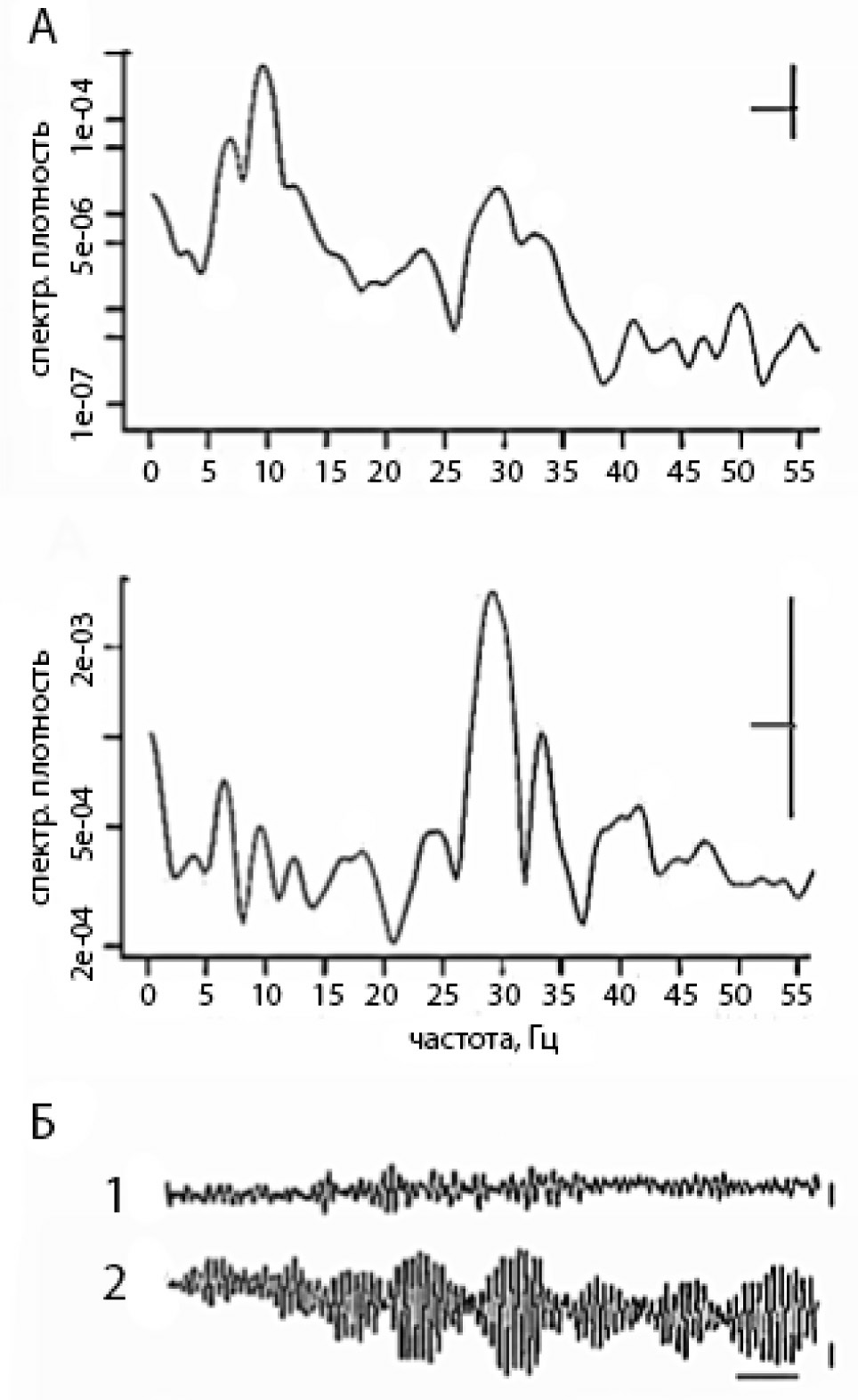
Рис.1. Частотные характеристики электрических осцилляций в двух локусах цианобактериальной
пленки — низкой и повышенной активности: А — периодограммы активностей, показанных на Б.
По оси абсцисс — частота в Гц, по оси ординат — спектральная плотность в условных единицах.
Горизонтальная черта — полоса пропускания, вертикальная черта — 95% доверительный интервал.
Б — полевые потенциалы, зарегистрированные одновременно в спокойном и активном локусе
биопленки Oscillatoria terebriformis. 1 — осцилляции спокойной зоны, 2 — осцилляции в активной
зоне. Калибровка: 20 мкВ, 300 мс.
Уровень синхронизации полевых потенциалов между областями определяли при помощи коэффициентов частотной и частотно-временной когерентности (рис. 2Б, 2В). Высокий уровень синхронизации, при котором коэффициент частотной когерентности достигает максимального значения, характерен для активных зон (рис. 2Б); график частотновременной когерентности показывает, что высокий уровень синхронизации сохраняется в течение довольно длительного времени — в приведенном случае 6 с (не исключено, что и дольше: анализируемый интервал ограничен временем наблюдения). Иначе выглядят показатели, демонстрирующие синхронность осцилляций между активным и спокойным локусом (рис. 2В): достижение максимального значения коэффициента когерентности в этом случае является кратковременным, может располагаться как на низких, так и на высоких частотах, что отражает график частотно-временной когерентности.
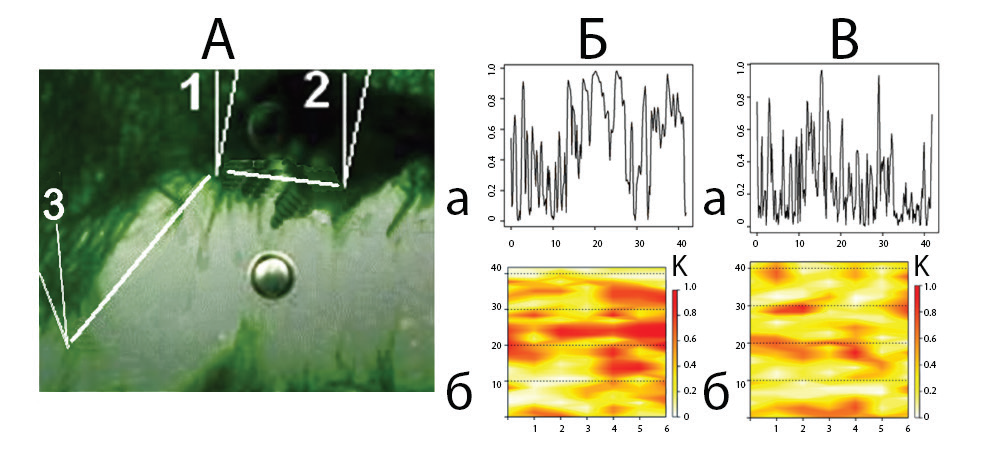
Рис.2. Взаимодействие между локусами цианобактериальной пленки, индивидуумы которых в разной степени вовлечены в активное строительное поведение: А — расположение электродов;
Б — фазово-частотная (а) и частотно-временная (б) когерентность электрической активности элементов, расположенных в центре зоны активности (электроды 1 и 2); В — в зоне регистрации электродов (3) и (1). Обозначения для (а): ось абсцисс — частота в Гц, ось ординат — коэффициент когерентности; для (б): ось абсцисс — время в секундах, ось ординат — частота в Гц; К — коэффициент когерентности
Обсуждение
Опыты, выполненные на цианобактериях, показали, что синхронизированные электрические осцилляции могут быть объективными показателями, характеризующими активность микробной социальной структуры. Главный результат опытов показал, что для выполнения социально значимой задачи необходим высокий уровень синхронизации электрической активности, инициируемой членами сообщества.
Идея о том, что синхронность колебаний биопотенциалов относится к числу электрографически выраженных явлений, благоприятных для функционирования элементов биологической системы, известна с 50-ых гг. прошлого века [Ливанов М.Н, 1972]. Основным методом исследования синхронности электрической активности различных участков биологического вещества является когерентный анализ. Например, в опытах на людях найдено, что при взаимодействии участников во время решения задачи для достижения общей цели осцилляторная активность определенных областей мозга синхронизируется и ее вспышки ассоциируются с действиями партнеров [Funane, 2011; Hu Yi, Hu, 2017]. Результаты охватывают объективные показатели в виде регистрации электрически выраженных событий головного мозга человека и животных, движений глаз и словесного отчета испытуемых при решении когнитивных задач, слушании музыки или текстов [Зотов, 2017; Bhattacharya, 2001; Kelong Lu, Ning, 2019; Kingsbury, 2019]. Эта активность по поводу достижения цели проходит на фоне синхронизированных процессов определенных областей мозга у каждого участника и между участниками [Snyder, 2018] и зависит от уровня их подготовки к определенному виду деятельности [Canolty, 2010]. Коэффициент когерентности тем выше, чем выше уровень синхронизации электрических потенциалов.
Результаты выполненных нами измерений позволяют заключить, что синхронизация активностей при решении задач, требующих совместных действий, необходима не только для многоклеточных существ, но и для микроорганизмов, которые решают проблемы, объединив усилия тысяч индивидов. Опыты показали, что уровень синхронизации, измеряемый коэффициентом когерентности, дает возможность оценить эффективность поведения у живых существ, занимающих первые строчки в истории развития жизни, в данном случае у прокариот — цианобактерий Oscillatoria terebriformis (рис. 2 Б).
Бактерии существуют в сообществах и координируют свое поведение для выполнения специфических функций. Техника генетики и молекулярной биохимии в соединении с микроскопной визуализацией показала, что развитие биопленки является хорошо регулируемым процессом, в котором бактерии интегрируются в сообщества посредством внутренней и внешней сигнализации. Сложность создания биопленки заставляет предполагать, что это способ развития, в котором изменение формы и функции играет ведущую роль в жизненном цикле бактерий. Создание сообщества требует координированных действий участников, между которыми происходит коммуникация [Олескин, 2009; Романова, 2006; Ben-Jacob, 1998; Masi, 2015]. Межклеточные контакты, представленные разнообразными экстрацеллюлярными структурами — микрофибриллами, шишковидными выступами, эвагинатами клеточных стенок, капсулами, — отражают генетически детерминированную закономерность развития микробных популяций как саморегулирующихся многоклеточных систем [Новик, 1995].
Таким образом, уровни синхронизации электрической активности у цианобактерий Oscillatoria terebriformis обеспечивают эффективность поведения этих существ: когерентность сигналов выше для локусов, в которых усилия элементов сообщества целенаправленны. Цианобактериальная пленка не полностью охвачена «строительными работами»: выделены области, от членов которых требуется наиболее активное участие. Это означает, что происходит дифференциация пространства, разделение его на пострадавшие зоны и зоны благополучные для жизни сообщества. Очевидно, происходит функциональная специализация: в работу по восстановлению пленки вовлекаются ближайшие к месту повреждения цианобактериальные нити. О феномене «разделения труда» в микробном социуме свидетельствуют факты электрофизиологических опытов [Греченко, 2019] и данные микробиологических исследований [Рыбальченко, 2003]. Предположение о специализированных членах сообщества опирается на результаты работ по электронной микроскопии — в частности, в ряде работ показана морфологическая гетерогенность микробных популяций, установлены закономерности в изменении структуры микробных сообществ на разных этапах развития, проявляющиеся в изменении соотношения различных типов клеток: физиологически активных, покоящихся, автолизированных и инволюционных [Рыбальченко, 2003; Von Bronk]. Гетерогенность популяции представляет собой результат реализации адаптивного потенциала, изначально свойственного конкретному микроорганизму. Иными словами, она является инструментом раскрытия новых приспособительных возможностей одного и того же бактериального генома [Магданова, 2013]. Полученные результаты совпадают с результатами работ, выполненных на людях и высокоорганизованных животных [Ливанов М.Н, 1972; Шарова, 1980; Ahn, 2015; Snyder, 2018]. Опыты на живых существах разного эволюционного уровня показывают, что истоки избирательной пространственной синхронизации, в результате которой устанавливаются функциональные отношения между разнородными зонами современных биологических систем — зонами мозга, участками биопленки и др., — возникли в жизни одних из самых ранних организмов нашей планеты. Актуализация этих функциональных отношений в форме поведения определенного типа является результатом интеграции деятельности этих зон.