Введение
На сегодняшний день актуальным направлением электрофизиологических исследований мозга является изучение функциональной связи ритмической активности тета-диапазона с формированием и реализацией поведения и его психологическими характеристиками.
Значительное внимание исследователей привлекает тета-ритм. Под тета-ритмом обычно понимают медленноволновую активность мозга с частотой 4—8 Гц и амплитудой до 200 мкВ. В качестве основных источников генерации тета-активности принято считать структуры медиального септума и диагонального пучка Брока, а также гиппокамп и прилежащие структуры, которые анатомически и функционально тесно связаны с префронтальными и фронтальными областями коры. Показано, что интеграция активности указанных структур осуществляется в значительной степени на основе тета-активности [19; 24].
Традиционно осцилляторную активность на частоте тета-ритма соотносят с различными психическими процессами. В частности, принято считать, что увеличение спектральной мощности тета-ритма отражает рабочее напряжение индивида и готовность к осуществлению деятельности [4; 13]. Известно, что усиление спектральной составляющей тета-ритма в ЭЭГ характерно для состояния эмоционального напряжения, когнитивной нагрузки и коррелирует с процессами памяти и обучения [13; 30; 35; 42], а также отражает такие неспецифические факторы, как общий уровень внимания [10]. В ряде работ модуляции внимания связываются с осцилляциями на частоте тета-ритма (4—8 Гц) [43; 44].
Упомянутая выше ассоциация активности в тета-диапазоне ЭЭГ с процессами формирования и поддержания памяти специально обсуждается в большом количестве исследований [26; 30]. Так, например, в задачах на эпизодическую память выявлена связь увеличения мощности тета-ритма с успешностью запоминания [37]. Показано, что стимуляция на частоте тета-ритма (методом транскраниальной стимуляции) определенных зон коры, прежде всего префронтальных и теменных, приводит к синхронизации ритмической активности нейронов на соответствующей частоте и увеличивает результативность выполнения мнестических заданий [18; 22; 45]. Тета-ритм также рассматривается как необходимый компонент механизмов контроля, обслуживающей процессы рабочей памяти [40]. В работах П.В. Симонова изменения тета активности коры связывались в основном с эмоциональной сферой [14]. При этом в генезе эмоциональных реакций и организации целенаправленного поведения ведущая роль отводилась четырем мозговым структурам: передним отделам новой коры, гиппокампу, миндалине и гипоталамусу. В дальнейшем было показано, что в эмоциональной ситуации миндалевидное тело и гиппокамп оказывают взаимное влияние, при этом обе структуры участвуют в мозговом обеспечении рабочей памяти [39].
Таким образом, из приведенных выше данных можно сделать вывод, что тета-ритм сопровождает процессы памяти, внимания, обучения новым навыкам и тесно связан с эмоциональным напряжением. Однако при рассмотрении вышеизложенных проявлений тета-ритма остается открытым вопрос, почему разнообразные процессы, необходимые для выполнения любой целенаправленной деятельности, эмоции разной модальности, сопровождаются тета-ритмом, который, как уже отмечалось, относят к медленным ритмам. По-видимому, причину этого следует искать в работе единого базового механизма, который является необходимой составляющей всех этих процессов.
В этой связи особый интерес представляет анализ активности мозга, в том числе ритмической, с позиций системно-эволюционного подхода [2; 17]. Согласно ему, процесс индивидуального развития сопряжен с выработкой новых типов адаптивного поведения, способствующих достижению полезного приспособительного результата — формированием функциональных систем в процессах системогенеза [2; 17]. Выработка нового типа адаптации начинается с рассогласования между данным и потребным соотношением индивида со средой и невозможностью его устранения с помощью имеющихся в структуре индивидуального опыта систем. Таким образом, индивидуальное развитие представляет собой последовательность системогенезов, в ходе которых происходит накопление и усложнение индивидуального опыта [2; 17]. С позиций системно-эволюционного подхода, функциональные системы, сформированные на ранних этапах (приобретения опыта) онтогенеза, обеспечивают относительно простые способы взаимодействия индивида с окружающей средой. Такие системы могут быть названы низкодифференцированными. По мере развития индивида его взаимодействие со средой усложняется и степень дифференцированности его индивидуального опыта возрастает [3]. При этом необходимо отметить, что в любом поведении достижение результатов обеспечивается за счет одновременной активности элементов опыта разного «возраста» и степени дифференциации [2; 3; 7; 17]. Также показано, что в ситуации рассогласования, подразумевающей необходимость формирования нового типа поведенческой адаптации, наблюдается временное упрощение взаимодействия индивида со средой, сопровождающееся увеличением эмоциональности и вклада низкодифференцированных систем в осуществление поведения — регрессия [1; 5]. Приведены эмпирические и теоретические аргументы в пользу того, что временная (обратимая) системная дедифференциация — начальный этап, необходимый для эффективного формирования нового навыка [2]. Положительный эффект временной дедифференциации обусловлен упрощением формирования нового домена опыта за счет подавления высокодифференцированных систем других доменов и минимизации необходимости в согласовании этих элементов опыта друг с другом в процессе научения [2].
Известно, что дельта— и тета-ритмы относятся к более филогенетически древним типам ритмической активности [31]. Дельта-ритм характерен для мозга рептилий, тета-ритм — для низших млекопитающих [31; 38]. Онтогенетическое созревание ЭЭГ человека является рекапитуляцией филогенетического развития. ЭЭГ новорожденных детей в основном представлена низкочастотными осцилляциями [12; 16]. По мере развития происходят изменения ЭЭГ, состоящие в увеличении спектральной мощности высокочастотных ритмов [8; 12; 15]. Принято считать, что активность дельта-ритма коррелирует с выраженностью витальных и биологических мотиваций, в то время как тета-ритм сопровождает адаптивные перестройки организма в ситуации изменившихся условий среды [20; 21; 23; 27] и эмоциональные процессы [9; 14]. В частности, показано увеличение спектральной мощности тета-ритма при воспроизведении наиболее ярких и образных автобиографических воспоминаний [32], а также увеличение спектральной мощности тета-ритма при рассматривании эмоциональных изображений лиц [9; 25; 33].
В рамках системно-эволюционного подхода можно предположить, что увеличение спектральной мощности низкочастотных ритмов отражает актуализацию низкодифференцированных систем. Изменение состава актуализированных систем в ходе реализации поведения и степень их дифференцированности можно оценивать по изменению показателей вариабельности сердечного ритма, сопряженных с эмоциональными характеристиками поведения [6]. В то же время спектральная мощность тета-ритма, соотносящаяся с процессами научения и повышенной эмоциональностью, может выступать в качестве еще одного признака дедифференциации. В пользу этого положения свидетельствуют данные, демонстрирующие, что введение этанола, которое приводит к блокированию относительно новых систем вновь приобретенных навыков [2; 7], сопровождается увеличением вклада тета-ритма в общемозговую активность [36].
С учетом сказанного выше нами выдвинута гипотеза, согласно которой увеличение спектральной мощности тета-ритма отражает регрессию, находится во взаимосвязи с эффективностью формируемого поведения и наблюдается на этапе выработки алгоритма решения новой задачи, т. е. на начальном этапе формирования навыка.
Целью настоящей работы явилась проверка выдвинутой выше гипотезы при исследовании изменения спектральной мощности тета-диапазона ЭЭГ на этапах подготовки и решения новой когнитивной задачи у участников эксперимента, демонстрирующих разную результативность ее выполнения.
Материалы и методы
В исследовании на основе добровольного информированного согласия участвовали 32 студента, юноши в возрасте 18—21 года, правши с нормальной или скорректированной до нормальной остротой зрения. Исследования одобрены локальным этическим комитетом ФГАОУ ВО Первого МГМУ имени И.М. Сеченова Минздрава России (Сеченовский университет) № 02-20 от 05.02.2020. Участник эксперимента находился в затемненной комнате в отсутствие посторонних раздражителей, сидя на расстоянии 50 см от экрана монитора. Использовался монитор Acer G276 с диагональю 27 дюймов.
В качестве модели когнитивной деятельности использовалась «n-back» задача (задача непрерывного выполнения), разработанная психологом В.К. Кирхнером [29]. Задача используется в нейрофизиологических исследованиях, а также в психологии для оценки и развития рабочей памяти, логического мышления и способности к концентрации внимания. В нашем варианте суть задания состояла в том, что человеку один за другим на экране в центре монитора предъявлялся ряд геометрических фигур (треугольник, квадрат или круг, зеленого, синего или красного цвета). Геометрические фигуры демонстрировались на сером фоне. В задачу участника эксперимента входило запоминать эти фигуры и указать (кнопкой мыши), встречалась ли представляемая в данный момент фигура два предъявления назад (рис. 1).
У каждого участника эксперимента регистрировали фоновую ЭЭГ в исходном состоянии. На первом этапе работы участник эксперимента получал подробную инструкцию о выполнении предстоящего задания и на конкретных примерах последовательностей геометрических фигур (тренировочной задачи) обучался выполнять его. Первый этап обучения каждого участника проходил при контроле и помощи исследователя и продолжался до безошибочного выполнения тренировочной задачи. Сразу после завершения первого этапа обучения у каждого участника эксперимента регистрировали ЭЭГ. На втором этапе обучения участники эксперимента самостоятельно выполняли тест. [Более детальное описание процедуры исследования представлено в работе: 10]. По результатам выполнения компьютерной задачи для каждого участника эксперимента вычисляли количество правильно указанных геометрических фигур (максимальное число правильных выборов составляло 10), которые, согласно инструкции, соответствовали фигуре, демонстрирующейся два предъявления назад.
Таким образом, схему исследования можно представить в следующем виде:
— ФОН — регистрация ЭЭГ в исходном состоянии при открытых глазах (60 с);
— ОБУЧЕНИЕ 1 — инструкция и обучение — выполнение участниками эксперимента теста «n-back» под контролем и с помощью исследователя (без регистрации ЭЭГ);
— ПЕРЕХОДНЫЙ ПЕРИОД — состояние ожидания предстоящей самостоятельной работы — 2-го этапа обучения. Регистрация ЭЭГ при открытых глазах (60 с).
— ОБУЧЕНИЕ 2 — самостоятельное выполнение участниками эксперимента теcта «n— back». Регистрация ЭЭГ при открытых глазах (130 с)
ЭЭГ регистрировали помощью электроэнцефалографа «Нейрон-спектр» (г. Иваново) монополярно по схеме «10—20» в затылочных (О2, О1), теменных (Р4, Р3), центральных (С4, С3), лобных (F4, F3) и височных (Т4, Т3) отведениях. Объединенные референтные электроды располагаются на мочках ушей. Полоса фильтрации составляет 0,5—70,0 Гц, постоянная времени — 0,32 с, режективный фильтр — 50 Гц. Импеданс был ниже 10 кОм для всех каналов. После регистрации все записи ЭЭГ были переведены в компьютерную систему анализа и топографического картирования электрической активности мозга «BRAINSYS» для Windows и обработаны с помощью аппаратно-программного комплекса «НЕЙРО-КМ» (ООО «Статокин», г. Москва). Артефакты исключали из анализируемой записи с использованием возможностей программного комплекса BRAINSYS. Спектральный анализ ЭЭГ проводили на основе быстрого преобразования Фурье (пакет программ BRAINSYS). Эпоха анализа составляла 4 сек, частота оцифровки — 200 Гц. Рассчитывали абсолютные значения спектральной мощности (СМ, мкВ2) общего диапазона тета-ритма ЭЭГ (4—7 Гц). Артефакты, возникающие при движении глаз, дифференцировали по их характерной форме и пространственному распределению и исключали из анализируемой записи с использованием возможностей программного комплекса BRAINGAM (в частности, процесса автоматического распознавания артефактов с визуальным контролем).
Для статистического анализа и представления данных использовали пакет программ «STATISTICA v.10.0». Данные представлены в виде [Me (Q1; Q3)]. Для проверки нормальности распределения переменных использовали критерий Шапиро—Уилка. Сравнение групп проводили при помощи критерия Краскела—Уоллеса как непараметрического аналога дисперсионного анализа для несвязанных выборок (Kruskal—Walles ANOVA). Изменения показателей в разных ситуациях у одной группы участников эксперимента оценивали с использованием критерия Фридмана («Friedman ANOVA by Ranks»), являющегося непараметрическим аналогом дисперсионного анализа повторных измерений. Апостериорный анализ проводили с использованием критерия Манна—Уитни (Mann—Whitney U test) для несвязанных групп. Динамику показателей у одной группы участников эксперимента оценивали по Т-критерию Вилкоксона (Wilcoxon test). Проводили ранговый корреляционный анализ по Спирмену. Гипотеза о достоверности различий принималась при р < 0,05.
Результаты
Анализ результатов выполнения теста «n-back» показал, что по группе участников эксперимента в целом число правильных ответов в тесте варьировало от 3 до 10, составив в среднем 7,47±0,36. По результату теста «n-back» выделены две группы испытуемых. Первую группу составили участники эксперимента, которые показали 8—10 правильных ответов из 10 возможных (n=12). Число правильных ответов в этой группе составило 9,50±0,15. Во вторую группу (n=9) вошли участники эксперимента, у которых число правильных ответов было не более 6 из 10 возможных. Результат участников эксперимента второй группы составил 4,77±0,36. Значения СМ тета-ритма ЭЭГ у участников эксперимента выделенных групп на этапах обследования представлены на рис. 2 и 3.
В исходном состоянии (ФОН) достоверных различий СМ тета-ритма между участниками эксперимента выделенных групп во всех анализируемых отведениях ЭЭГ не обнаружено. На этапе «ПЕРЕХОДНЫЙ ПЕРИОД» СМ тета-ритма была выше у участников эксперимента первой группы в правом и левом затылочных (О2, р=0,049; О1, р=0,004), в правом и левом теменных (Р4, р=0,007; Р3, р=0,003), правом и левом центральных (С4, р=0,030; С3, р=0,040), левом лобном (F3, р=0,030) отведениях ЭЭГ. Выявленные различия обусловлены тем, что у участников эксперимента первой группы имело место увеличение СМ тета-ритма уже на этапе «ПЕРЕХОДНЫЙ ПЕРИОД» по сравнению с исходным состоянием, достоверно в левых затылочной, теменной, центральной и височной областях коры (рис. 2). Наряду с этим, у участников эксперимента второй группы значимых изменений СМ тета-диапазона на этапе «ПЕРЕХОДНЫЙ ПЕРИОД» относительно этапа «ФОН» не зарегистрировано.
У участников эксперимента первой группы по сравнению с участниками эксперимента второй группы на этапе «ОБУЧЕНИЕ 2» СМ тета-ритма была выше в правом и левом затылочных (О2, р=0,040; О1, р=0,020) и правом теменном (Р4, р=0,016) отведениях ЭЭГ. На этапе «ОБУЧЕНИЕ 2» относительно этапа «ФОН» увеличение СМ тета-ритма имело место только в левых фронтальном и теменном отведениях ЭЭГ. В то же время значимых изменений СМ тета-ритма на этапе «ОБУЧЕНИЕ 2» относительно этапа «ПЕРЕХОДНЫЙ ПЕРИОД» не зарегистрировано (рис. 2).
У участников эксперимента второй группы на этапе «ОБУЧЕНИЕ 2» по сравнению с этапом «ФОН» имела место более высокая СМ тета-ритма в правом и левом теменных, центральных и фронтальных отведениях ЭЭГ. На этапе «ОБУЧЕНИЕ» относительно этапа «ПЕРЕХОДНЫЙ ПЕРИОД» значимое увеличение СМ тета-ритма зарегистрировано в левом теменном, правом и левом центральных и фронтальных отведениях (рис. 3).
Таким образом, у участников эксперимента первой группы на этапе «ПЕРЕХОДНЫЙ ПЕРИОД» наблюдалось увеличение спектральной мощности тета-ритма относительно исходного состояния в отведениях ЭЭГ левого полушария. В отличие от них у участников эксперимента второй группы значимое увеличение спектральной мощности тета-ритма наблюдалось только в период самостоятельного выполнения задания.
Обсуждение
В настоящем исследовании были выделены две группы участников эксперимента с различной результативностью теста «n-back». У участников эксперимента, которые достигли более высокого результата теста, спектральная мощность биопотенциалов тета-диапазона увеличивалась относительно исходного состояния на этапе подготовки к самостоятельному выполнению задания и сохранялась на протяжении всего периода «ОБУЧЕНИЕ 2». В отличие от них у менее результативных участников эксперимента увеличение мощности тета-ритма ЭЭГ относительно исходного состояния и этапа «ПЕРЕХОДНЫЙ ПЕРИОД» имело место только на этапе «ОБУЧЕНИЕ 2».
Обращает на себя внимание тот факт, что у более результативных участников эксперимента увеличение спектральной мощности тета-ритма наблюдалось не на этапе самостоятельного выполнения теста, как показано в многочисленных исследованиях, а в период ожидания самостоятельной работы, в связи с выработкой алгоритма решения предстоящей задачи — первого этапа обучения. В работах П.В. Симонова приводятся данные о связи тета-ритма со степенью сформированности навыка: суммарное напряжение тета-ритма в ЭЭГ уменьшалось по мере стабилизации двигательного навыка у животных. При этом затруднения в реализации двигательного акта приводили к повторному нарастанию тета-ритма [14]. В работах Laukka et al. было показано увеличение вклада тета-ритма в общемозговую активность в период обучения выполнению задачи [35]. Также в работах Lang et al. отмечалось увеличение спектральной плотности тета-ритма у участников эксперимента в период обучения выполнению визуально-моторного теста по сравнению с исходным состоянием [34]. Важно отметить, что большее увеличение спектральной плотности тета-ритма наблюдалось у более результативных участников эксперимента, что согласуется с полученными нами данными.
Мы полагаем, что выявленное в настоящем исследовании увеличение медленноволновой активности в тета диапазоне на этапе «ПЕРЕХОДНЫЙ ПЕРИОД» может отражать первый, «регрессионный», этап научения, характеризующийся увеличенным вкладом низкодифференцированных систем в системную организацию поведения.
Одно из функциональных значений временной дедифференциации в процессе формирования нового опыта или усовершенствования уже имеющегося заключается в повышении эффективности и ускорении научения [2]. Таким образом, можно предположить, что более высокая результативность у участников эксперимента первой группы, демонстрировавших повышение спектральной мощности тета-ритма после первого этапа обучения, связана с эффективным формированием нового поведения, направленного на выполнение предлагаемой в рамках тестирования когнитивной задачи. В свою очередь, увеличение спектральной мощности тета-ритма во время второго этапа обучения — самостоятельного выполнения теста, наблюдавшееся у участников эксперимента второй группы, по всей видимости, является следствием более поздней дедифференциации, обусловленной рассогласованием между необходимостью справиться с заданием и отсутствием сформированной модели поведения.
В целом, представленные в литературных источниках результаты исследований предоставляют противоречивую информацию о направленности изменений показателей тета-ритма в различных зонах коры при выполнении когнитивных задач [29; 42]. В подобных исследованиях описывается сложная структура механизмов, через работу которых реализуются процессы внимания и памяти, активность которых может существенно меняться в зависимости от специфики выполняемой задачи. В то же время мы предполагаем, что все процессы, в той или иной степени сопровождающиеся увеличением спектральной мощности тета-ритма, соответствуют процессам, которые характеризуют те или иные аспекты поведения, разворачивающегося при временной дедифференциации в ходе формирования новой адаптации.
Сопоставление показателей повышения спектральной мощности тета-ритма и регрессии, обоснованное не только теоретически, но и полученными данными, требует, однако, дополнительных исследований. Имеющиеся данные относительно изменения ритмической составляющей мозговой активности в ходе индивидуального развития [16; 31], а также психические процессы, сопровождающиеся увеличением тета-активности, в частности, процессы внимания и памяти, эмоциональные переживания [9; 25; 32; 33], наряду с явлениями, которые описываются при системном анализе регрессии, дают дополнительные основания для проведения таких параллелей и соответствий.
Как было отмечено выше, с позиций системно-эволюционного подхода, формирование нового навыка выполнения задания начинается с дедифференциации: возрастания вклада низкодифференцированных систем в обеспечение поведения [2; 5]. Согласно данным настоящего исследования, у более результативных участников эксперимента была в большей степени выражена временная дедифференциация, которая сопровождалась увеличением вклада тета-ритма, уже на этапе, предшествующем самостоятельному выполнению теста. По-видимому, данный процесс способствовал более эффективному системогенезу и получению ими высокого результата. Наряду с этим, у менее результативных участников эксперимента увеличение спектральной мощности тета-ритма на этапе самостоятельного выполнения теста было связано с рассогласованием, обусловленным недостаточной сформированностью навыка решения поставленной когнитивной задачи и более поздним развертыванием дедифференциации.
Заключение
По результату выполнения теста «n-back» были выделены две группы участников эксперимента. В исходном состоянии по спектральной мощности тета-ритма участники эксперимента этих групп не различались. После первого этапа обучения выполнению задачи спектральная мощность тета-ритма была выше в затылочных, теменных, центральных и левой фронтальной областях коры у успешных участников эксперимента. В основе таких различий было увеличение спектральной мощности тета-ритма относительно исходного состояния в ряде отведений ЭЭГ только у результативных испытуемых. В отличие от них, у менее результативных участников эксперимента значимое увеличение спектральной мощности тета-ритма (по сравнению с исходным состоянием и состоянием после первого этапа обучения) имело место позже: лишь на втором этапе обучения.
Увеличение спектральной мощности тета-активности в период обучения может быть рассмотрено как «регрессионный» этап системогенеза, сопровождающийся увеличением вклада низкодифференцированных систем в осуществление нового поведения и обусловливающий сравнительно более высокую эффективность последующей деятельности.
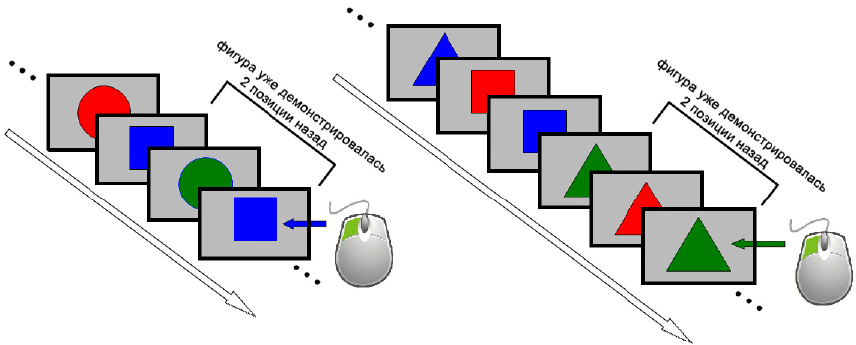
Рис. 1. Примеры последовательностей появления геометрических фигур на экране монитора теста n-back. Отмечены фигуры (уже демонстрировавшиеся два предъявления назад), при появлении которых необходимо нажать левую кнопку мыши
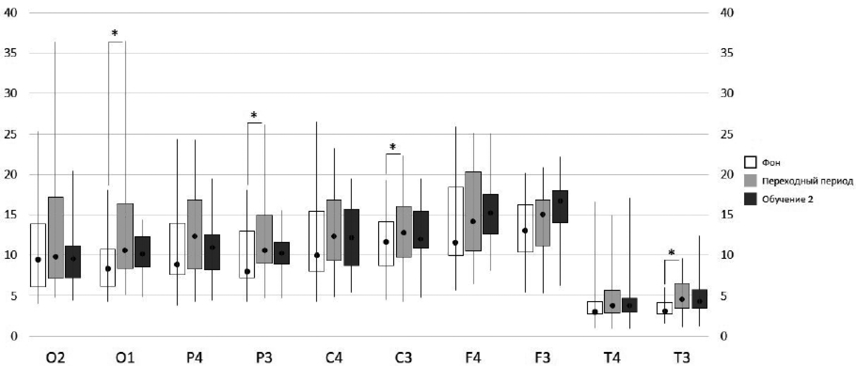
Рис. 2. Спектральная мощность тета-ритма у участников эксперимента первой группы в исходном состоянии (ФОН, белый столбик), сразу после получения инструкции и обучения (ПЕРЕХОДНЫЙ ПЕРИОД, серый столбик) и на этапе выполнения теста (ОБУЧЕНИЕ 2, темно-серый столбик); «*» — достоверные различия (р<0.05) в значениях мощности тета-диапазона ЭЭГ на разных этапах
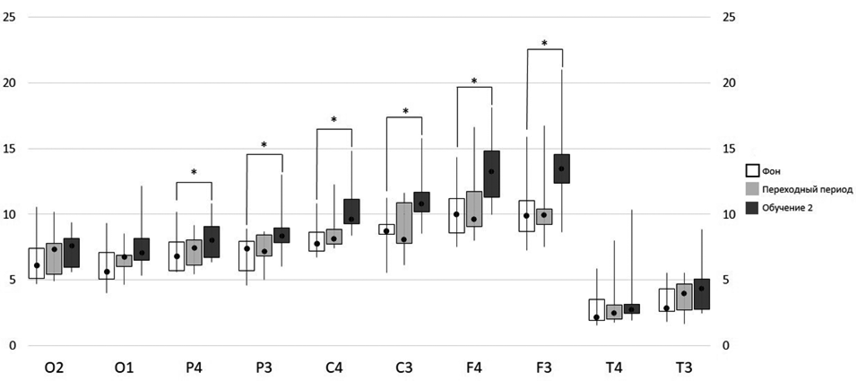
Рис. 3. Спектральная мощность тета-ритма у участников эксперимента второй группы в исходном состоянии (ФОН, белый столбик), сразу после получения инструкции и обучения (ПЕРЕХОДНЫЙ ПЕРИОД, серый столбик) и на этапе выполнения теста (ОБУЧЕНИЕ 2, темно-серый столбик); «*» — достоверные различия (р<0.05) в значениях мощности тета-диапазона ЭЭГ на разных этапах