До недавнего времени проблема локализации речевых зон в коре мозга человека казалась практически решенной. Еще в конце XIX в. были выявлены две важнейшие речевые зоны: зона, отвечающая за восприятие обращенной к человеку речи, — зона Вернике, расположенная в височной коре, и зона, связанная со способностью порождать речевое высказывание, — зона Брока, находящаяся в нижнелобных отделах. Как правило, у праворуких испытуемых обе зоны расположены в левом полушарии [Geschwind, 1970]. Однако оказалось, что исключения из этого «правила» встречаются достаточно часто. Согласно данным Бенсона, чье исследование признается одним из наиболее репрезентативных, среди праворуких пациентов левополушарная локализация речи была выявлена примерно у 90 % [Benson, 1985]. В остальных случаях имело место «атипичное» распределение речевой функции по полушариям. У леворуких пациентов это распределение еще более размыто.
Первоначально локализация в коре зон Брока и Вернике была определена на основании данных, полученных в клинике органических поражений мозга [Broca, 1861]; [Wernicke, 1874]. Впоследствии «золотым клиническим стандартом» локализации этих речевых зон в том или ином полушарии стал инвазивный тест, предложенный в 50-х годах прошлого века канадским физиологом Дж. А. Вада [Wada, 1997]. Поочередная кратковременная инактивация полушарий коры мозга с помощью введения в соответствующую каротидную артерию наркотизирующего вещества позволяет определить «доминантное по речи» полушарие у конкретного пациента и таким образом прогнозировать функциональные выпадения и резервы полушарий большого мозга пациента после проведения нейрохирургического вмешательства. Данный метод может применяться только с целью защиты речевых зон от возможных повреждений во время нейрохирургической операции по удалению опухоли, абсцесса или резекции резистентного эпилептического очага. Метод категорически не пригоден для применения в исследовательской практике. Достаточно высокий процент осложнений, возникающих при проведении данного теста (риск кровоизлияния составляет 10—15 %), создает необходимость изобретения новых методов локализации речевых зон в коре мозга человека и для клинических целей. Другая проблема этой методики заключается в ее недостаточной пространственной точности: она позволяет определить латерализацию речевой функции, но не дает никакой возможности локализовать речевые области внутри полушария, затрудняя таким образом предоперационный прогноз хирургического вмешательства.
В последние десятилетия многие исследователи стали использовать современные методы нейровизуализации: функциональную магнитно-резонансную томографию (фМРТ) и позитронно-эмиссионную томографию (ПЭТ), чтобы разрешить проблему неинвазивной локализации когнитивных функций (в том числе речевой) в коре мозга человека. Основной вывод из этих исследований заключался в том, что речевые стимулы, по сравнению с неречевыми, вызывают более массированную активацию в верхней височной извилине в обоих полушариях, а феномен левополушарного доминирования обнаруживается в умеренной степени и только на групповом уровне [Demonet, 1992]; [Hickok, 2004]; [Vouloumanos, 2001]; [Zatorre, 1996].
Неопределенность локализации в значительной степени связана с выбором методов исследования. Восприятие речи человеком — чрезвычайно быстрый процесс: смысл слова извлекается из акустических характеристик речевого сигнала в течение долей секунды. Поэтому особенно на ранних стадиях работа отдельных мозговых структур, участвующих в его обработке, совершается почти мгновенно — за десятые доли секунды [Shtyrov, 2007а]. ПЭТ и фМРТ позволяют визуализировать медленные (секундные) изменения активности мозга на основании параметров интенсивности кровотока. Таким образом, эти методы являются недостаточно информативными с точки зрения получения данных о мозговых основах речевых прессов, поскольку не способны уловить кратковременную активацию нейронных популяций. Лишь технологии, обладающие возможностью непосредственной регистрации электрической нейронной активности, такие как электро- и магнитоэнцефалография (ЭЭГ, МЭГ), могут обеспечить точную информацию о временной динамике.
Методика МЭГ основана на измерении минимальных (порядка 10-13 — 10-15 тесла, T) магнитных полей и их градиентов, генерируемых синхронной активностью больших масс нейронов головного мозга. Это достигается с помощью сверхпроводимых сенсоров, расположенных в вакууме с температурой, близкой к абсолютному нулю (температура ~4 по Кельвину поддерживается с помощью жидкогогелия). Дальнейшая обработка полученных данных включает сложные компьютерные алгоритмы, с помощью которых сначала выборочно усиливается и определяется та небольшая часть записанного сигнала, которая исходит собственно от мозга. Этот сигнал необходимо очистить от мощного общего электромагнитного фонового шума. После того как это сделано, необходимо идентифицировать внутренние источники мозговой активности на основе внешней записи. Эта обратная задача является в данном случае по определению некорректной, поскольку любые внешние показатели могут быть потенциально объяснены с помощью более чем одной конфигурации внутренних источников. В контексте психофизиологического эксперимента ее решение возможно лишь при наложении ограничений, исходящих из природы экспериментальной задачи и знаний о мозговых анатомических структур и их функций.
Таким образом, задача функционального картирования определяется не только адекватным выбором метода измерения функциональной активности мозга, но и, в не меньшей степени, выбором методических подходов и экспериментальных парадигм, которые смогли бы максимально эффективно использовать технологию регистрации и обработки вызванной электро-магнитной активности мозга.
При выборе экспериментальной парадигмы необходимо учитывать, что анализ речевой информации сопровождается динамическими изменениями активности нейронных ансамблей, причем в доминантном по речи полушарии эти изменения значительно сильнее, чем в «неречевом» [Binder, 1997]; [Fiez, 1996]; [Giraud, 2001]; [Howard, 1992]; [Mummery, 1999]; [Narain, 2003]; [Scott, 2000]. Для успешного понимания речи мозгу требуется мгновенно расшифровать информацию на самых разных уровнях [Norris, 2000]: он должен успеть определить звуки, классифицировать их как фонемы (т. е. именно как речевые звуки), разбить на морфемы и слова, установить между ними грамматические связи и использовать их для того, чтобы построить целостную структуру фразы и понять ее смысл, в конце концов встроив его в общий ситуативный контекст.
Слуховой речевой сигнал обладает комплексом физических характеристик, таких как частотные характеристики, длительность пауз, амплитуда и скорость изменения частотного состава. Речевые зоны мозга человека должны обработать акустические особенности слухового речевого стимула, чтобы приступить к его фонематическому, а затем и семантическому анализу. Возникает вопрос, в каком временном окне мозг начинает обрабатывать речевой сигнал именно как речь, а не как любой другой сложный слуховой стимул? Только после нахождения ответа на этот вопрос можно будет переходить к проблеме локализации зон мозга, занятых в этом процессе.
Экспериментальные данные показывают, что семантический анализ произнесенного слова начинается в мозге примерно через 200—300 мсек после начала его звучания [Connolly, 1995]; [Helenius, 2002]; многие исследователи предлагают и намного более ранний интервал: 50—150 мс [Shtyrov, 2007]; [MacGregor, 2012]. Следовательно, к этому времени мозг либо уже осуществил фонематическую обработку слухоречевого стимула — задача, осуществляемая нейросетями в зоне Вернике [Liebenthal, 2005], либо фонематические процессы идут параллельно с остальным анализом речи на более высоком уровне. У праворуких людей при тяжелом повреждении зоны Вернике в левом полушарии теряется способность воспринимать речь, а речевые фонемы воспринимаются ими как шум [Geschwind, 1970]. Таким образом, можно предположить, что речевые зоны включаются в обработку уже в течение первых 200 мс звучания слова, и нам необходимо знать, как и что именно происходит в эту пятую долю секунды.
Теоретическая психолингвистика выделяет четыре уровня обработки речи: (1) фонологическая обработка — идентификация фонетических/фонологических единиц в звуковом потоке, (2) лексическая — соотнесение последовательности услышанных фонем с «умственным словарем», (3) семантическая — определение смысла речевого стимула и (4) синтаксическая — анализ грамматической информации, содержащейся в высказывании.
До сих пор в литературе существуют противоположные взгляды на вопрос, как упорядочены процессы, происходящие на разных уровнях. Одни исследователи предполагают, что обработка речевых сигналов происходит последовательно, путем передачи информации от одного уровня обработки к другому [Friederici, 2002]. Другие отстаивают гипотезу о параллельной обработке речевых сигналов на этих уровнях [Hagoort, 2008]; [Marslen-Wilson, 1987]; [Shtyrov, 2007а].
В пользу второго предположения существует ряд экспериментальных подтверждений [Shtyrov, 2010]; [Shtyrov, 2005]; [Shtyrov, 2007] Данные нейроанатомических и функциональных исследований говорят о том, что передача информации между кортиевым органом и корой больших полушарий по самым быстрым путям занимает не более 10—20 мс [Eldredge, 1971], дальнейшие 15—20 мс уходят на то, чтобы передать сигнал из основных слуховых зон в нижние отделы лобной доли и в височные области [Shtyrov, 2007]. Следовательно, существуют сильные нейробиологические доводы в пользу того, что как минимумпервичное извлечение важной лингвистической информации из речи (если не интеграция ее в более широкий контекст) может осуществляться уже через ~50 мс после начала обработки.
Единственным способом получения прямых экспериментальных доказательств или опровержений этих гипотез является метод МЭГ, обладающий, как уже было указано выше, временным разрешением на уровне миллисекунд и, в комбинации со структурной МРТ, миллиметровым пространственным разрешением.
Подобные исследования проводились с использованием так называемой оддболл-парадигмы (odd7ball paradigm). В этой парадигме частые (стандартные) слуховые стимулы подаются вперемешку с редкими (девиантными) стимулами. Разница в ответе слуховой коры мозга на редкие стимулы по сравнению со стандартными получила название негативности рассогласования — mismatch negativity (MMN) [Alho, 1995]; [Naatanen, 1992]. Обычно MMN рассматривают как отражение активации нейронных сетей кратковременной слуховой памяти, локализация которых в мозге зависит от обрабатываемой слуховой информации. Неудивительно поэтому, что MMN на слуховые речевые стимулы ведет себя иначе, чем на неречевые [Aulanko, 1993]; [Phillips, 2000]; [Shtyrov, 2000]
Для решения задачи латерализации и локализации речевых зон важна следующая особенность возникновения MMN на речевые стимулы. Именно речевые звуки (фонемы и слова) сопровождаются сильной левосторонней асимметрией MMN, тогда как равные им по сложности и интенсивности неречевые слуховые стимулы вызывают одинаковую активацию слуховой коры в обоих полушарий [Gootjes, 1999]; [Vihla, 2003]. Более того, ответ MMN на речевые фонемы в левом полушарии задержан, по сравнению с ответом на неречевые стимулы, уравненные с фонемами по сложности физических характеристик [Eulitz, 1995]; [Tiitinen, 1999]. Более того, локализация паттерна MMN на слова с разным смыслом отражает семантические свойства слов, что дополнительно указывает на чувствительность этого ответа к лингвистической информации [Pulvermuller, 2005]. В связи с этими данными можно предположить, что локализация MMN на речевые стимулы в мозге испытуемого может способствовать определению его речевых зон, включающихся в работу уже на этапе обработки фонематической информации, содержащейся в прозвучавшем слове [Parviainen, 2005]. Многократно показано, что выделяемый в вызванной речевыми стимулами магнитной активности мозга компонент MMN локализуется в первую очередь в области верхневисочной извилины левого полушария [Naatanen, 1997]; [Shestakova, 2002]; [Shestakova, 2004]; [Shtyrov, 2005]. Более того, характерные для этого мозгового ответа высокая временная точность, высокий уровень соотношения сигнала к шуму и автоматизм генерации (методика не требует внимания испытуемых к речи) позволяют предложить его в качестве перспективного метода картирования речевых зон как у здоровых испытуемых, так и у пациентов, ожидающих нейрохирургическое операционное вмешательство.
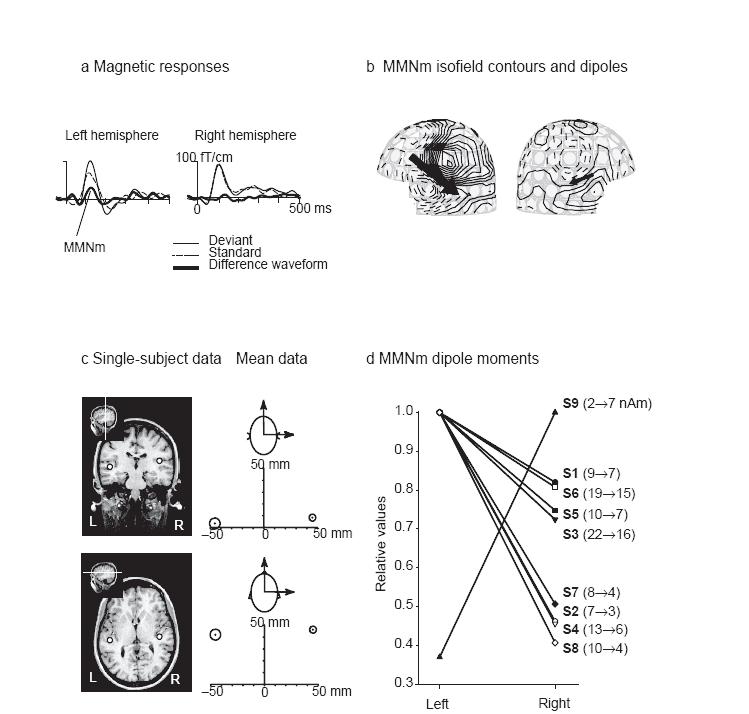
Рис. Пример МЭГответов испытуемого на речевые стимулы. [Aulanko, 1993] а) представляет собой типичный пример ответов, зарегистрированных на градиометрах (типе сенсоров), представляющих ответы обоих полушарий. На (в) представлены изопотенциальные карты топографии силовых линий магнитного поля с шагом в 8 fT). На (c) части иллюстрации показана идея использования индивидуальной МР томограммы испытуемого для более точной локализации нейронального ответа в височной области; (d) часть представляет собой анализ межполушарной асимметрии по результатам МЭГ картирования для каждого испытуемого. (Адаптировано из Shestakova et al., 2004)
МЭГ метод является также чрезвычайно перспективным для изучения принципов организации лингвистических следов памяти. Бесконечные попытки идентификации уникальной зоны «хранилища слов» в мозге не увенчались успехом, и сегодня в целом принято, что контуры памяти на слова имеют свойство «распределенности» [Mesulam, 1990]. Возникает вопрос о принципах формирования таких распределенных ансамблей. Существует предположение, что их формирование происходит по принципу ассоциации в процессе взаимного усиления связей между различными (даже удаленными друг от друга) областями коры мозга. Эти связи образуются в то время, когда человек выучивает действия, слова или понятия в сочетании со словами, которые используются для их описания [Pulvermuller, 2010]. Референциальные значения являются неотрывной частью семантики слова, и мы считаем, что в мозге они реализуются посредством реализации ассоциативного научения распределенных нейронных популяций по принципу Хебба — при одновременной активации пре- и постсинаптических областей синаптическая передача усиливается[Morris, 1999].
Если лексические репрезентации представлены как распределенные в коре нейронные контуры, а действия, которые означают эти слова, соматотопически отражаются в моторных зонах мозга [Penfield, 1950], из этого следует конкретное предположение, что слова, обозначающие действия различных частей тела (например, рука или нога), также будут вызывать активацию соответствующих зон моторной коры. Это утверждение подтверждается данными ЭЭГ, МЭГ и фМРТ, что соматотопически специализированные зоны моторной коры активируются в ответ на классы слов, «кодирующих» действие соответствующих частей тела [Hauk, 2008]; [Pulvermuller, 2005]; [Shtyrov, 2004]. МЭГ-данные особенно примечательны тем, что они показывают динамику распределенной активности вовлеченных участков коры с высокой временной и пространственной точностью [Boulenger, 2012].
Таким образом, за последнее десятилетие стало очевидным, что использование МЭГ-технологии для исследования мозговой активности, связанной с обработкой речи у взрослых и детей, открывает возможность получения качественно новых знаний о процессах восприятия и порождения речи, а также о механизмах освоения языка (родного и иностранного). Совмещение данных МЭГ с результатами МРТ сканирования головного мозга позволят практически впервые разработать методы неинвазивного функционального картирования мозга с целью локализации речевых зон и определения степени их латерализации.
В Центре нейрокогнитивных исследований (МЭГ-центре) МГППУ в настоящий момент осуществляется проект по разработке методов и экспериментальных парадигм для определения латерализации процессов восприятия речевых сигналов на слух и процессов кратковременной слухоречевой памяти.