Введение
Освоение субъектом новых навыков и опыта при столкновении с ситуацией новизны способствует также более успешному воспроизведению того опыта, который был сформирован непосредственно после ситуации новизны. Например, было показано, что уже знакомые изображения (лица или пейзажи) распознаются лучше, если их повторное предъявление сопровождается новыми картинками, по сравнению с аналогичными ситуациями, когда в сопровождение включены только знакомые картинки [Bunzeck N, Duzel, 2006]. При этом новизна и позднее формируемый опыт могут принадлежать доменам, напрямую не связанным, так, например, результаты эксперимента с предъявлением фотографий жизненных ситуаций (ранее не предъявляемых), за которым следовала задача категоризации слов, свидетельствуют о более успешном их воспроизведении на следующий после исследования день [Fenker D.B, 2008]. На аналогичную взаимосвязь указывают и данные эксперимента, в котором испытуемым предлагалось проводить активное исследование нового виртуального пространства — чем выше испытуемые оценивали реализм своих ощущений от новой виртуальной среды, тем лучше воспроизводились предъявленные после первой процедуры слова [Schomaker, 2014]. Новизна также может способствовать улучшению свойств памяти: животные в модели пассивного избегания электрического тока демонстрировали такую память, если обучались этому после активного исследования новой обстановки [Moncada D, Viola, 2007].
Однако необходимо отметить, что новизна сама по себе не всегда оказывает такой положительный эффект на воспроизводимость памяти [Biel, 2019]. Результаты исследования Байла и Банзека [Biel, 2019] указывают на отсутствие положительной динамики показателей распознавания слов, выученных непосредственно до или после просмотра видеофрагментов, в ситуации пассивного просмотра новых видеофрагментов без активного выполнения какой-либо задачи. Можно предположить, что во всех этих работах речь идет о феномене переноса как характеристики взаимодействий между элементами опыта [Созинов, 2013]. Объяснение неоднозначности получаемых результатов требует обращения не к анализу эффектов памяти, а к изучению механизмов запоминания и воспроизведения после ситуации новизны, поскольку необходимо установить, что именно изменяется при формировании нового опыта на фоне новизны, что позднее может приводить к изменениям в воспроизведении какого-либо навыка. Так, было показано, что помещение мыши в частично совпадающую с ситуацией обучения обстановку (т. е. напоминающую о ранее приобретенном опыте, но и частично обладающую новизной) приводит к индукции экспрессии раннего гена c-fos и реконсолидации [Amelchenko, 2012]. Известно, что белок Fos (продукт экспрессии гена c-fos) маркирует нейроны, претерпевающие нейрогенетические изменения в связи с текущим научением [Анохин, 1997]. Основная задача настоящего исследования заключалась в изучении особенностей выполнения различных поведенческих актов и выраженности нейрогенетических изменений в головном мозге у крыс в ходе научения пищедобывательному навыку при воспроизведении ранее приобретенного в ситуации новизны навыка.
Методы
Исследование проводилось на самцах крыс линии Long-Evans, массой 200—380 грамм, возраст 5—19 месяцев (n=13) (в эксперимент были отобраны животные двух возрастных групп: 5 месяцев и 19,5±1,5 месяцев, см. раздел «Результаты»). В течение всего времени проведения эксперимента животные содержались в своих домашних клетках размером 48х37х20 см. С момента начала обучения инструментальным навыкам животные экспериментальных групп находились на питьевой или/и пищевой депривации. Потеря веса за весь период обучения не превысила 10—12%.
В исследовании использовалось несколько экспериментальных клеток для формирования разных навыков. Тест открытого поля и тест на предпочтение объектов проводился в клетке № 1 размером 50х50 см с тремя белыми и одной прозрачной стенкой. Во время теста на предпочтение объектов в эту обстановку на пол прикрепляли две разные пластиковые игрушки размером около 5 см. Инструментальному питьевому поведению крыс обучали в другом помещении, где находилась белая квадратная клетка № 2 размером 50x50 см, передняя стенка которой также была прозрачной. В правом углу передней стенки находилась поилка, а в левом углу рядом с передней стенкой — рычаг, при касании которого с помощью вибриссной подушки в поилку подавалась вода. «Вибриссная» педаль длиной 3—4 сантиметра и шириной 1 сантиметр располагалась на уровне головы животного. Вне клетки находилась кнопка экспериментатора, нажатие которой позволяло экспериментатору подавать воду животному. Инструментальное пищедобывательное поведение животные выполняли в клетке № 3 размером 73x46x67 см, которая содержала две автоматические кормушки, расположенные в разных углах у передней стенки клетки. Две педали, нажатие на которые приводило к подаче кормушек, находились в противоположных углах у дальней стенки клетки. Вне клетки находилась кнопка экспериментатора, нажатие которой позволяло подавать кормушку с пищей в любое время, независимо от поведения крыс. Поведение животных во всех случаях регистрировалось при помощи цифровой видеокамеры.
В первые два дня для измерения общей активности животных помещали в открытое поле на 15 минут. Далее (на 3-й день) оценивалось предпочтение животными нового объекта. В этой задаче крыс на 5 минут помещали в клетку № 1, где находились две пластиковые игрушки. Регистрировалось общее время обследования каждого из объектов. Через 5 минут крыс вынимали, а одну из игрушек заменяли новой, закрепленной на том же месте, что и предыдущая. Аналогичную процедуру повторяли через 60 минут и через 24 часа после первого знакомства животного с новыми объектами, каждый раз заменяя «старый» объект на новый. Каждая сессия тестирования длилась 5 минут. Процедура данного тестирования, включая длительность самого теста и интервалы между разными сессиями, соответствовала процедуре, предложенной в исследовании Ennaceur, Delacour [Ennaceur A, Delacour, 1988]. Сразу после завершения теста на предпочтение крысам вводили частичную питьевую депривацию (с доступом к воде в течение 30 минут ежедневно) и начинали обучать инструментальному питьевому поведению. Обучение было разделено на отдельные этапы, каждому из которых животные обучались в течение одной ежедневной 30-минутной сессии. В первый день крысы получали порцию воды за подход к поилке. Во второй день животных обучали отворачиваться от поилки в сторону рычага. На третий день вода подавалась за отход от поилки к середине стенки по направлению к рычагу. На четвертый день крыс обучали подходу к рычагу. На пятый день обучения инструментальному питьевому поведению крысам подавалась вода, когда они проводили вибриссной подушкой по краю рычага. В последующие пять дней происходило закрепление полученного навыка и знакомство с пищедобывательной клеткой (животные помещались в эту экспериментальную клетку на 5 минут в течение пяти дней сразу после завершения сессии «вибриссного» поведения, при этом в кормушках не было пищи). В последний день часть животных (n=7) помещались на пять минут в ту экспериментальную клетку, где проводился тест на предпочтение объектов, при этом один объект перемещался на новое место относительно того, где он находился при последнем помещении крыс в эту обстановку [подробнее см.: 19]. Таким образом, моделировалась актуализация имеющегося опыта в условиях новизны (группа «актуализация»). Сразу после окончания данного теста этих крыс в течение 30 минут обучали в другой клетке (и другом помещении) пищедобывательному инструментальному поведению нажатия на педаль. С животными группы «активный контроль» производились все те же манипуляции (тест открытого поля, тест на предпочтение объектов, питьевое инструментальное обучение, пищедобывательное обучение) за исключением актуализации имеющегося опыта перед формированием пище- добывательного поведения (животных не тестировали на предпочтение объектов непосредственно перед пищедобывательным обучением).
Спустя 75 минут после окончания обучения пищедобывательному навыку животных усыпляли ингаляционным наркозом (эфиром). Сразу после этого проводили декапитацию, извлекали мозг и сразу же замораживали его в парах жидкого азота. Выявление нейрогенетических изменений в мозге осуществляли по продукту экспрессии одного из ранних генов белку c-Fos [Анохин, 1997]. Для этого на криостатных микропрепаратах срезов (толщиной 20 мкм) мозга проводили иммунногистохимические реакции в соответствии со стандартным протоколом: микропрепараты срезов мозга фиксировали в течение 15 минут в 4% растворе параформальдегида при температуре 4 градуса Цельсия, отмывались три раза однократным раствором фосфатного буфера (PBS), для снижения неспецифического окрашивания помещались в 0,3% раствор перекиси водорода на 30 минут, отмывались три раза 1*PBS, инкубировались с первичными антителами, разведенными 1 к 2000 (sc-52 rabbit, Santa Cruz Biotechnology) в течение 15 часов, отмывались пять раз ^PBS с добавлением Тритона, инкубировались со вторичными антителами (REAGENT KIT Anti-Rabbit, Vector), отмывались 1*TBS, инкубировались с авидин-стрептовидиновым комплексом в течение 1 часа, отмывались 1*PBS, помещались в раствор DAB (раствор 3,3’ — диаминобензидина с содержанием 0,3% перекиси водорода) и отмывались водой. После чего срезы проводили через батарею восходящих спиртов (70%, 96%, 100% по 3 минуты в каждом), а затем помещали в ксилол и заключали под покровные стекла. Оценку числа Fos-положительных клеток проводили на оцифрованных срезах мозга.
Далее осуществлялся анализ нейрогенетических изменений в бочонковом поле соматосенсорной коры, активность нейронов которого опосредует «вибриссное поведение» [например: 22; 9] и в ретросплениальной коре, где, как было ранее показано, обнаруживается большое число нейронов, специализированных относительно данного пищедобывательного поведения [например: 5; 2; 29]. Для каждого животного проводилось усреднение показателей по всем срезам. Для анализа изображений срезов мозга использовали программу ImageProPlus (“Media Cybernetics”, США).
Видеозаписи открытого поля и инструментального пищедобывательного поведения обрабатывались с помощью программы RealTimer (http://www.openscience.ru) и Trackvit [Коршунов, 2014]. Для инструментального пищевого поведения регистрировались: нажатия левой педали; нажатия правой педали; залезания в левую кормушку; залезания в правую кормушку; груминг; столбик; заход в левый дальний угол; заход в правый дальний угол; заход в левый ближний угол; заход в правый ближний угол. Для открытого поля регистрировались следующие поведенческие акты: столбик с опорой на стенку; столбик без опоры на стенку; груминг; чистка шерсти; обнюхивание; пересечения квадрата в центре поля; пересечения квадрата на периферии поля; заход в дальнюю левую часть; заход в дальнюю правую часть; заход в ближнюю левую часть; заход в ближнюю правую часть. Данные поведенческие акты регистрировались по количеству реализаций, по общему времени реализации, и также высчитывалось средние время каждого поведенческого акта. Статистический анализ характеристик поведенческих актов и нейрогенетического обеспечения формирования нового навыка проводили при помощи программы STATISTICA 6.0 и StatPlus.
Результаты
В результате серии последовательных научений животные формировали несколько групп компонентов опыта, связанных с нахождением в обстановке открытого поля (экспериментальная клетка № 1), взаимодействием с непищевыми объектами в открытом поле, вибриссо-опосредованным питьевым навыком в экспериментальной клетке № 2 и пище- добывательным навыком нажатия на педаль в экспериментальной клетке № 3. При этом нейрогенетические изменения в мозге оценивали только после формирования последнего навыка в экспериментальной клетке № 3.
Животные двух возрастных групп (см. раздел «Методы») не отличались по общеповеденческим характеристикам в обстановке открытого поля, а также по скорости перемещения и длине пройденного пути при обучении пищедобывательному навыку в клетке № 3 (критерий Манна—Уитни, p > 0,05 во всех случаях), поэтому для дальнейшего анализа крысы разного возраста были объединены в одну группу. Результаты анализа особенностей выполнения животными теста на предпочтение нового объекта описаны нами ранее [Кузина, 2018].
Было обнаружено, что после формирования пищедобывательного навыка нажатия на педаль в экспериментальной клетке № 3 число Fos-положительных нейронов в ретросплениальной коре не различалось между группами «актуализация» и «активный контроль» для обоих полушарий (критерий Манна—Уитни, z = 0,54 иz = 1,56 соответственно: р > 0,05; 11=6; п=5). Здесь и далее оценку нейрогенетических изменений производили отдельно для контралатерального и ипсилатерального полушарий (по отношению к используемой животным в питьевом навыке вибриссной подушке). Данные по плотности Fos-положительных нейронов в обоих полушариях ретросплениальной коры и бочонковом поле соматосенсорной коры представлены в табл. 1.
Таблица 1
Плотность Fos-положительных нейронов в ретросплениальной и соматосенсорной коре в группах «актуализация» и «активный контроль» (число нейронов в 1 мм2)
|
Группы |
Ретросплениальная кора |
Бочковое поле соматосенсорной коры |
||
|
Ипсилатеральное полушарие |
Контралатеральное полушарие |
Ипсилатеральное полушарие |
Контралатеральное полушарие |
|
|
Актуализация |
155 (114—270) |
106 (37—171) |
109 (109—110) |
50 (38—99) |
|
Активный |
88 |
94 |
51 |
57 |
|
контроль |
(86—128) |
(71—106) |
(48—70) |
(—59) |
Примечание: в скобках даны перцентили, 25% и 75% соответственно.
Оказалось, что число Fos-положительных нейронов между группами «актуализация» и «активный контроль» не различалось в ретросплениальной коре (рис. 1). Достоверные различия были обнаружены только для бочонковых полей соматосенсорной коры ипсилатерального полушария (критерий Манна—Уитни, z = 2,4; p = 0,01; n=5) (рис. 2). При этом пищедобывательное поведение животных групп «актуализация» и «активный контроль» достоверно не различалось (табл. 2).

Рис. 1. Индуцированные обучением пищедобывательному навыку нейрогенетические изменения (по белку Fos) в ретросплениальной коре в контралатеральном (левый график) и ипсилатеральном (правый график) полушариях по отношению к вибриссной подушке, использованной ранее в питьевом навыке
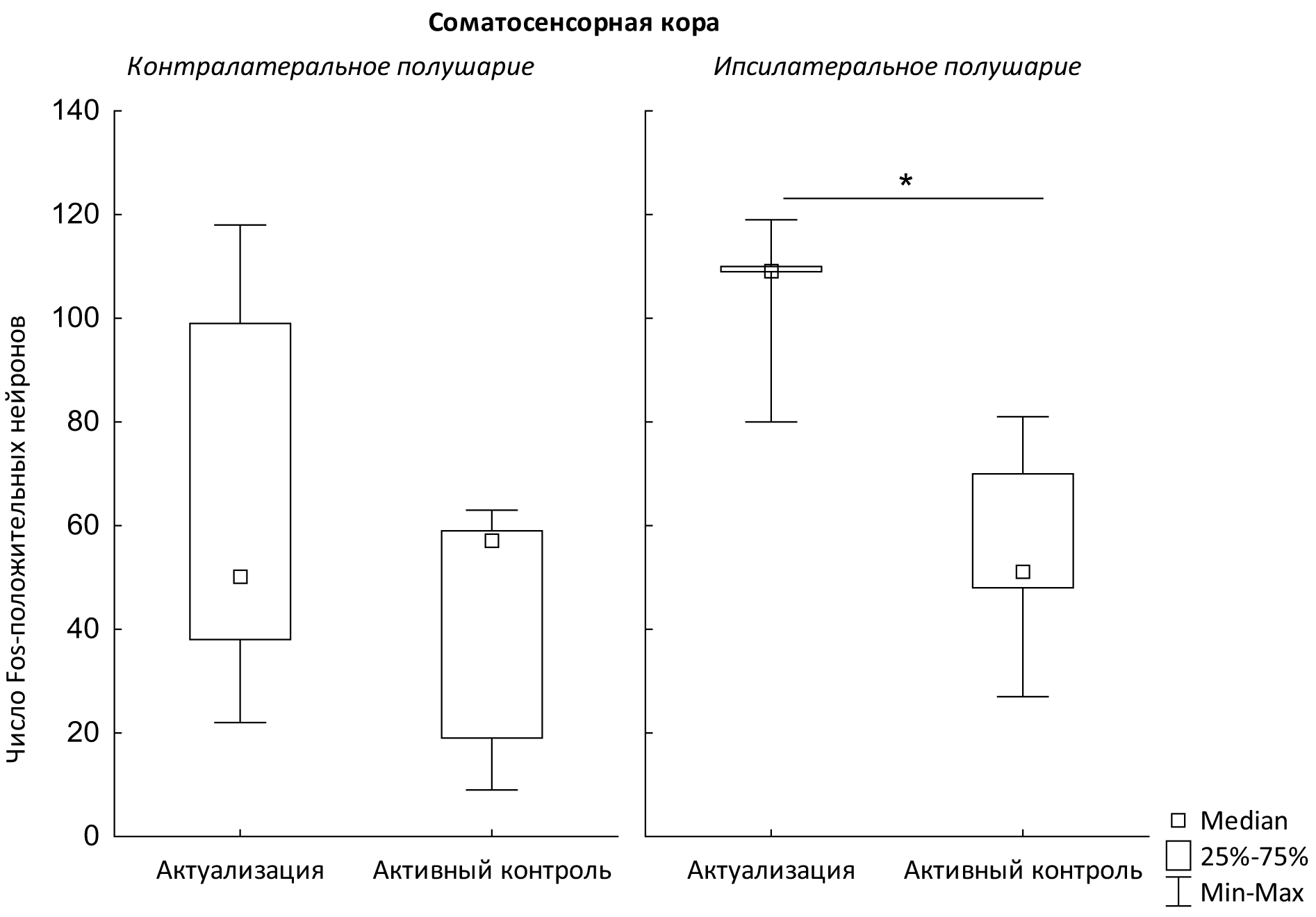
Таблица 2
Основные параметры (медианы и 1— 3 квартили) поведения животных групп «актуализация» и «активный контроль» при приобретении пищедобывательного навыка
|
Группы |
Время нахождения в зоне эффективных кормушек, % |
Время нахождения в зоне неэффективных кормушек, % |
Время в зоне эффективной педали, % |
Время в зоне эффективной педали, % |
Средняя длительность стоек,сек |
Суммарная длина пути, см |
|
Актуализация |
61,6 (54,83-74,97) |
11,73 (10,7-16,06) |
21 (12,73-24,5) |
3,3 (2,4-7,8) |
1,31 (0,75-1,65) |
14167 (9200-17458) |
|
Активный контроль |
71 (66,06-76,6) |
9,41 (8,47-13,06) |
12,55 (11,76-19,97) |
3,5 (2,83-4,43) |
1,8 (1,72-2) |
8203 (7743-8739) |
Например, как у животных группы «актуализация», так и у животных группы «активный контроль» наблюдался достоверно более высокий процент времени нахождения в зоне эффективных кормушек по сравнению в временем, проведенным в зоне неэффективных кормушек (критерий Вилкоксона, z = 2,37; p = 0,018 и z = 2,2; p = 0,028; n=7; n=6 соответственно), что свидетельствует об одинаково успешном прохождении этапа нахождения источника пищи в процессе обучения в обеих группах крыс. Также животные данных групп не различались по времени, проведенному в зоне эффективной педали (критерий Манна- Уитни, z = 1; p > 0,05; n=7; n=6). По другим актам, специфическим для выполнения пи- щедобывательного поведения, также не было обнаружено достоверных различий. Таким образом, кратковременная актуализация ранее сформированного опыта в новом контексте перед формированием нового элемента опыта не приводила к изменению «эффективности» научения, оцениваемой с помощью выбранных параметров. В то же время две экспериментальные группы животных различались по особенностям выполнения поведения, которое являлось «неспецифическим» [Ковалев, 2019] для формируемого пищедобывательного навыка. Так, средняя длительность выполнения стоек у животных группы «актуализация» была достоверно меньше, чем у животных группы «активный контроль» (критерий Манна-Уитни, z = -2,5; p = 0,012; n=7; n=6). У крыс группы «актуализация» суммарная длина пути при обучении нажатию на педаль была значимо больше, чем в группе «активного контроля» (критерий Манна-Уитни, z = 2,07; p = 0,038; n=7; n=6). Причем разница не обнаруживалась между группами в первом десятиминутном периоде (критерий Манна-Уитни, z = 1,35; p > 0,05; n=7; n=6) и во втором (критерий Манна-Уитни, z = 1,64; p > 0,05; n=7; n=6), но становилась достоверной в третьем периоде (критерий Манна-Уитни, z = 2,35; p = 0,0184; n=7; n=6). В то же время, в группе «актуализация» не было достоверных различий по пройденному пути между всеми десятиминутными периодами (критерий Вилкоксона, p > 0,05 во всех случаях, n=7), а в группе «активный контроль» достоверные изменения были обнаружены только между первым и вторым периодом (критерий Вилкоксона, z = 2,2; p = 0,027, n=6). Не было обнаружено достоверных различий по скорости перемещения (табл. 3) за все время сессии у животных между группой «актуализация» и группой «активный контроль» (критерий Манна-Уитни, z = 1,215; p > 0,05; n=7; n=6), однако в третьем периоде такие различия присутствовали: у животных группы «актуализация» скорость перемещения оказалась выше (критерий Манна-Уитни, z = 2,145; p = 0,0318: n=7; n=6). При этом у животных группы «актуализация» скорость не менялась между первым и вторым десятиминутным периодом, а также между вторым и третьим (критерий Вилкоксона, z = 1,26 и z = 1,35 соответственно; p > 0,05 во всех случаях; n=7). А у животных группы «активный контроль» скорость достоверно увеличивалась только от первого периода ко второму (критерий Вилкоксона, z = 2,2; p = 0,027; n=6). На основании полученных результатов можно предположить, что введение новизны или рассогласования имеющегося опыта с текущей ситуацией перед обучением новому поведению в ситуации новизны привело к ускорению распознавания обстановки, где проходило обучение инструментальному пищедобывательному поведению, как неопасной. Можно отметить также, что в исследовании особенностей формирования установки на обучение у другого вида животных (обезьян-гамадрилов) было обнаружено, что от экспериментальной активности животного зависела только вероятность нахождения правильного решения на первых этапах обучения, но не степень обученности, т. е. общая успешность формирования нового поведения [Аникаев, 2020], что согласуется с полученными в настоящей работе результатами об отсутствии различий в успешности обучения крыс с групп «актуализация» и «активный контроль».
Таблица 3
Средняя скорость перемещения (см/сек) животных групп «актуализация»
и «активный контроль» при приобретении пищедобывательного навыка.
В скобках даны перцентили, 25% и 75%, соответственно
|
Группы |
Скорость перемещения в первом 10-минутном периоде |
Скорость перемещения во втором 10-минутном периоде |
Скорость перемещения в третьем 10-минутном периоде |
Средняя скорость перемещения за все время сессии |
|
Актуализация |
7 (4,7—9,3) |
8,6 (5,2—10,7) |
8,6 (7—11,3) |
8,17 (5,3—10,7) |
|
Активный контроль |
5,6 (5—6) |
6,05 (6—6,2) |
5,65 (5—6,3) |
5,68 (5,67—6) |
Обсуждение
Таким образом, было установлено, что актуализация имеющегося опыта в ситуации новизны перед следующим обучением приводит к повышению общей активности животных, т. е. к изменению «неспецифического» поведения при формировании нового опыта, но не ускоряет само научение. При этом мозговое обеспечение формирования нового опыта зависит от этой предварительной актуализации ранее сформированного опыта, в частности, число Fos-положительных нейронов оказывается у таких животных больше. В свете полученных в настоящем исследовании результатов можно представить следующий сценарий формирования нового элемента индивидуального опыта.
В настоящее время в научной литературе все чаще приводятся аргументы в пользу того, что любое поведение формируется на основе актуализируемого в конкретной ситуации предшествующего опыта индивида [например: 27; 24]. С точки зрения селекционных теорий научения [например: 2; 12], в процессе приобретения нового опыта происходит формирование новой специализированной группы нейронов (из числа молчащих нейронов или из нейронов, образованных в процессе взрослого нейрогенеза), активность которых связана с выполнением приобретенного поведения. Однако системное описание научения на нейронном уровне включает в себя две группы связанных процессов: уже упомянутые процессы системной специализации и процессы аккомодационной реконсолидации, связанные с реактивацией имеющегося опыта [см., например: 13; 14]. Поскольку индивидам свойственно формировать обобщенную структуру знаний для разных форм деятельности, а не множество «локальных» для каждого нового навыка, процессы модификация структуры опыта животных при последовательном обучении серии разнородных навыков во многом перекрываются [Behrens, 2018; Jacklin, 2016]. В исследовании Jacklin и соавторов также было показано, что когда крысы обучаются различать новые объекты, активность таких структур мозга, как задняя теменная и периринальная кора, различается в зависимости от того, была ли ситуация обучения полностью или частично новой для животных [Jacklin, 2016]. Известно, что любое научение начинается с рассогласования между задачей в текущей для индивида ситуации и имеющимися у него компонентами индивидуального опыта, необходимыми для достижения требуемого результата [например: 1]. Таким рассогласованием является и новизна [Alexandrov, 2018]. Рассогласование между прогнозируемой (на основании филогенетического и онтогенетического опыта) и текущей ситуацией приводит к комплексу пробного поведения или ориентировочно-исследовательскому поведению. По-видимому, такие данные и были получены в нашем исследовании, только комплекс процессов, запущенных в одной ситуации, продолжал проявляться в последующем поведении.
Можно предположить, что и формирование новых специализаций нейронов, и реорганизация активности клеток, специализированных относительно ранее сформированных элементов (систем) опыта, маркируются нейрогенетическими изменениями [например: 10]. В наших экспериментах ранее было показано, что обучение пищедобывательному навыку вызывает экспрессию белка с-Fos в достоверно большем числе нейронов бочонкового поля у животных, обучавшихся предварительно инструментальному питьевому («вибриссному») навыку, чем в аналогичной области контрольных животных, предварительно обучавшихся неинструментальному питьевому навыку [Сварник, 2014]; это позволяет предположить, что активация экспрессии c-Fos при втором обучении происходила и в тех нейронах, которые уже являлись специализированными относительно первого, «вибриссного», навыка. Таким образом, в качестве первого этапа научения можно выделить запуск нейрогенетических изменений в нейронах систем уже имеющегося опыта, в силу каких-то причин актуализированного в настоящее время. Интересно, что в настоящем исследовании не обнаружилось достоверного увеличения числа Fos-положительных нейронов в контралатеральном бочонковом поле у животных обеих групп по сравнению с ипсилатеральным полушарием. Такой результат может быть объяснен использованием в исследовании дополнительных тестов распознавания непищевых объектов, которое также осуществлялось с помощью вибрисс. Тем не менее, полученные результаты показывают, что в ситуации дополнительного рассогласования (актуализация ранее сформированного опыта в ситуации новизны) было увеличено число нейронов, в которых наблюдается изменение экспрессии генов, по крайней мере, в исследованных корковых областях, что согласуется с описанными ранее данными о том, что именно ситуация рассогласования приводит к реконсолидации памяти [например: 26], а новизна связана с индукцией нейрогенетических изменений [например: 30].
Таким образом, на основании полученных данных можно высказать предположение, что актуализация ранее сформированного опыта перед новым обучением способствует формированию нового элемента опыта, но не в смысле ускорения формирования нового поведения, а в аспекте увеличения числа нейронов, меняющих экспрессию своих генов, что в дальнейшем может способствовать извлечению этого опыта и проявляться как «усиленная» память.